![]()
![]() The
possibility of a subjective level of neurological organization. The
functional representation of the subjective animal system of representation
makes it clear that any animals that happened to try out such a behavior guidance
system would be able to acquire an entire range of new powers for controlling
relevant conditions in a long period of gradual evolution. They would, in
other words, begin a new stage of animal evolution, refining the inherent
powers of spatial imagination and adapting it to all available ecological
niches. But in order to conclude that such an evolutionary stage is inevitable,
it must be possible for a radical random variation to try it out. There are
good reasons to believe that it is possible, but in the end, the most compelling
case depends on seeing how these functions are served by nervous mechanisms.
The
possibility of a subjective level of neurological organization. The
functional representation of the subjective animal system of representation
makes it clear that any animals that happened to try out such a behavior guidance
system would be able to acquire an entire range of new powers for controlling
relevant conditions in a long period of gradual evolution. They would, in
other words, begin a new stage of animal evolution, refining the inherent
powers of spatial imagination and adapting it to all available ecological
niches. But in order to conclude that such an evolutionary stage is inevitable,
it must be possible for a radical random variation to try it out. There are
good reasons to believe that it is possible, but in the end, the most compelling
case depends on seeing how these functions are served by nervous mechanisms.
It may seem that any neural mechanism complex enough to realize this functional description of the subjective animal system of representation would be too complex to be tried out as a radical random variation on the telesensory system by the multicellular biological behavior guidance system. The functional diagram indicates that it requires a higher level of neurological organization than telesensory animals. But since the mechanism of embryological development must get all its instructions from the structure of the nucleus and the chromosomes it contains, such a higher level may seem beyond the range of random variations being tried out, especially considering that more complex neural connections are needed for higher level systems to interact as an animal behavior guidance system.
This is the same kind of problem that we faced in explaining the eukaryotic and multicellular levels of biological organization. But there it required a new kind of biological behavior guidance system, and here a different kind of solution is required, for the same biological behavior guidance system is responsible for all the levels of neurological organization. The appropriate random variation must somehow occur in the multicellular biological behavior guidance system (the process of embryological development).
There are at least three reasons for believing that such a random variation is possible.
First, one reason was found in how deuterostome embryological development is different from proterostome development. With the neural tube divided into segments, the mechanism of embryological development can address entire 2-D arrays of neurons and determine them, as a whole, to migrate or send processes to other areas of the neural tube (or the body), and to make topologically ordered connections with 2-D arrays of neurons there. That means, as suggested earlier, that the units from which deuterostome development constructs the nervous system are already on a higher level of organization than in proterostome development — not to mention that the connections among such units can be made without interference with the development of other organs in the body, since the neural tube itself is separate from the rest of the body.
Second, it is not necessary for the mechanism of embryological development to prescribe all the neural connections involved in the subjective animal system of representation, because the three basic systems are laid out in such a way that most of the detailed connections they require can be acquired from the world by learning. Since the power of animal behavior comes from adapting it to spatial and spatio-temporal aspects of the world, it is possible to internalize the detailed structural causes of the brain mechanisms that serve that function from experience with the world. The world already has a spatial structure, and if the animal is protected and cared for during the first part of its life, its interactions with the world can internalize the structures needed to construct local images and maps of local images that can be explored by covert locomotion. Thus, much detailed structure can be supplied by the contained form of reproductive causation mentioned above, in which behavioral schemata evolve by reinforcement selection. This is the role of nurture as well as nature in neurological development.
Third, given how detailed neural connections can be internalized from the structure of the world by experience, that is, learning, all that is necessary is a random variation that would set up the basic structure on a higher level of neurological organization within which those detailed structures can be acquired. As it happens, there are two, connected kinds of changes in the embryological development of the non-mammalian vertebrate that do set up a higher level of neurological organization in the mammalian forebrain.
In order to give this final argument for the possibility of the subjective level of neurological organization, therefore, we must consider the structure of the mammalian brain.
Starting with the functional diagram of the subjective animal behavior guidance system, I will, first, simply identify the neural mechanisms in the mammalian brain that are responsible for each of the subsystems. This is to view the structure of the mammalian brain from a rather high altitude. It affords a general impression of how the mammalian brain works, and for those who are familiar with its structure, it will be possible to see how it explains the functioning of the mammalian brain.
Second, I will explain how this functional explanation of the structure of the mammalian brain would also explain the contained form of reproductive causation by which behavioral schemata evolve in the individual brain by reinforcement selection.
Finally, in order to show how the subjective level of neurological organization is possible, I will show how it could arise from two basic, yet simple changes in the non-mammalian vertebrate brain. That will involve a much more detailed description of how the functionally identified systems are realized in the mammalian brain.
![]() Neural
structure of the subsystems of the subjective animal behavior guidance system.
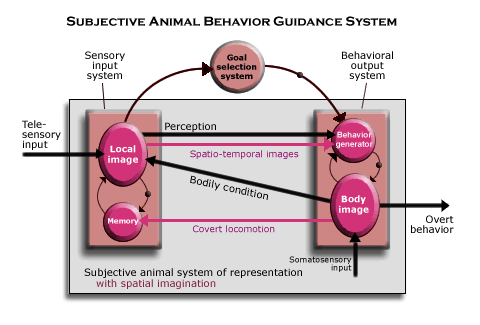
In the diagram of the subjective animal behavior guidance system, the
systems that were previously defined functionally are labeled by parts of
the nervous mechanism that serve their functions in the mammalian brain. (The
functional diagram is included for comparison.) The brain mechanisms involving
the neocortex are depicted as beveled buttons, while the thalamic nuclei projecting
to them are depicted as purple spheres. Notice that when we look more closely
at the detailed structure, the behavior generator itself is a complete
circuit of telesensory-level nervous mechanisms. That is because there are
two, distinct circuits within the behavioral output system, one that supplies
the behavioral schemata and generates covert behavior and the other that also
generates overt behavior. (Covert behavior needs only the ventral anterior
thalamic nucleus, whereas both it and the ventral lateral thalamic nucleus
are involved in generating overt behavior.) The part of the frontal neocortex
that contains the behavior schemata and where covert behavior is generate
is labeled “Sch.”)
Neural
structure of the subsystems of the subjective animal behavior guidance system.
In the diagram of the subjective animal behavior guidance system, the
systems that were previously defined functionally are labeled by parts of
the nervous mechanism that serve their functions in the mammalian brain. (The
functional diagram is included for comparison.) The brain mechanisms involving
the neocortex are depicted as beveled buttons, while the thalamic nuclei projecting
to them are depicted as purple spheres. Notice that when we look more closely
at the detailed structure, the behavior generator itself is a complete
circuit of telesensory-level nervous mechanisms. That is because there are
two, distinct circuits within the behavioral output system, one that supplies
the behavioral schemata and generates covert behavior and the other that also
generates overt behavior. (Covert behavior needs only the ventral anterior
thalamic nucleus, whereas both it and the ventral lateral thalamic nucleus
are involved in generating overt behavior.) The part of the frontal neocortex
that contains the behavior schemata and where covert behavior is generate
is labeled “Sch.”)
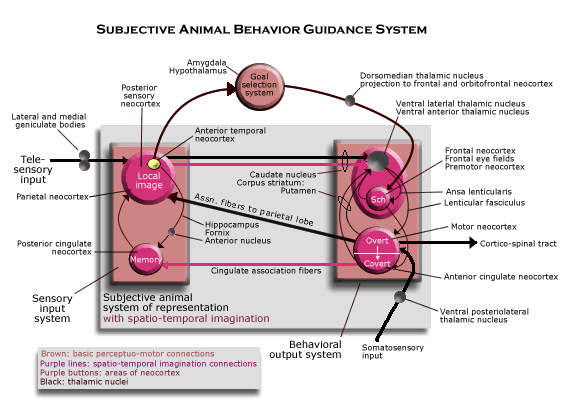
This key to the neurological interpretation of the functional diagram of the subjective animal behavior guidance system may be enough for those who are familiar with the structure of the mammalian brain to see the functional diagram as an explanation of how the mammalian brain works. But for those who are not familiar with the structure of the mammalian brain and want a briefer, more intuitive way of seeing how it realizes these functionally defined systems, the following diagrams and brief explanations may suffice.
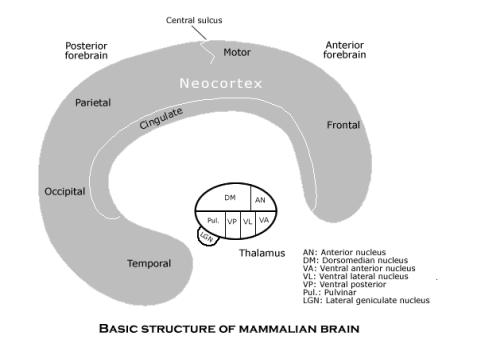
In the mammal, the entire animal behavior guidance system is located in the forebrain, and its basic structure is composed of two main units, the thalamus and the neocortex. The thalamus is at the end of a brain stem, just rostral to the midbrain, hindbrain and spinal chord, and it sends a massive projection to all parts the neocortex, which encloses the rest of the forebrain. (A projection is a set the axons from a 2-D array of neurons in one region to a 2-D array of neurons in another region, taking full advantage of the chordate’s way of constructing the nervous system in the neural tube.) The nuclei in the thalamus send their projections mostly to different parts of the brain (though the anterior temporal cortex receives none). There are also projections back from the neocortex to thalamic nuclei by which they interact in some way, but the main direction of influence is from the thalamus to the neocortex. These thalamic projections are 2-D arrays of neurons in which the information conveyed by the firing of any single neuron depends on the neuron's position in the 2-D array as a whole, and the thalamus apparently has the function of synchronizing the processing in all the parts of the neocortex.
There are three grand, distinct circuits of connections, each involving many such 2-D arrays, that pass though the thalamus and the neocortex back. That is, each circuit uses some part of the projection of many 2-D arrays from the thalamus to the neocortex, but each has a special way by which the neocortex completes the circuit of multiple 2-D projections by acting on those same thalamic nuclei (that is, distinct from the connections by which the neocortex reacts to the thalamus directly). Each such circuit is one of the three main subsystems of the animal behavior guidance system, with the circuits themselves on the subjective level of neurological organization. (See diagram of the basic structure of the mammalian brain.)
In the framework of this basic diagram, we can see how each of the functionally defined subsystems is realized by the mammalian brain. Consider first the sensory input system. Visual input from the eyes is relayed by the lateral geniculate nucleus (labeled "LGN") to the occipital (or striate) cortex where there are numerous copies of the entire image from the retina of the eye. Auditory input is not represented in the diagram, though it is handled in the same way, with auditory images in the superior temporal lobe. The inferior parietal neocortex is where the telesensory images are assembled as the local image (using association fibers from the motor areas, or body image, to supply information about the current bodily condition, also not depicted). The activity of the inferior parietal area in constructing the local image also depends on its interactions with a thalamic nucleus called the "pulvinar" or (labeled "Pul.," which receives input from the superior colliculus in the midbrain, the main visual image in the non-mammalian vertebrate brain).
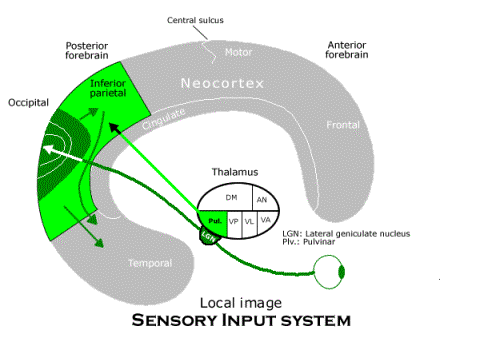
The memory is what gives the sensory input system a complete circuit from the thalamus through the neocortex back to the thalamus for another projection to the neocortex. The memory circuit receives information about the telesensory images and how they have been combined as the local image in the anterior temporal neocortex where the hippocampus is located. In fact, almost every region of neocortex sends association fibers (or chains of association fibers) toward the anterior temporal neocortex, where they are assembled for the hippocampus (in an adjacent region, called the “entorhinal cortex”). The hippocampus projects by way of the fornix to the anterior nucleus of the thalamus, though many of its fibers pass first to the mammillary body (located in the hypothalamus) which relays them to the anterior nucleus. The anterior nucleus projects to the entire cingulate gyrus, which is the extreme medial edge of neocortex adjacent to both anterior and posterior regions of neocortex. The cingulate neocortex has two way connections with neurons in adjacent areas of neocortex. That completes the memory circuit for the local image and its bundle of telesensory images, since those images are affected by their own projection through the memory circuit. And the function of this circuit is to form memory groups of neurons throughout the neocortex that are involved at some moment in the (synchronized) processing the image of some local scene. That is, by strengthening the synapses among the neurons involved throughout the neocortex, the whole group tends to fire when some crucial group of them fires for some reason, recalling the local image.
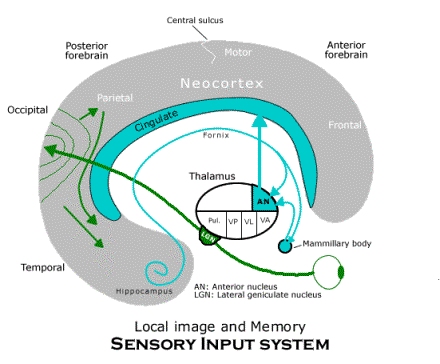
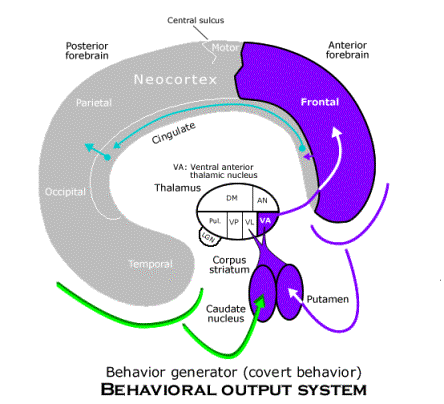
The behavior generator for covert behavior receives the local image (and its telesensory images) by way of the projection from the entire posterior neocortex to the caudate nucleus of the corpus striatum. That is the perception of the object in space that is used to guide behavior. The corpus striatum also receives a projection from the entire anterior neocortex, which includes both its perception of its own body and the behavioral schemata contained in the frontal neocortex. The corpus striatum can project to both the ventral lateral and ventral anterior nuclei of the thalamus, but when behavior is covert, the projection to the ventral lateral nucleus is suppressed and only the ventral anterior nucleus is affected. The ventral anterior nucleus projects to the entire frontal neocortex (except for the motor neocortex itself and supplementary neocortex). Thus, when behavior is covert, the premotor neocortex and behavioral schemata located in the frontal area are both also projected to the anterior cingulate gyrus, and since the anterior cingulate neocortex projects to the posterior neocortex and, indirectly, to the local image, covert locomotion can call up images from memory in sequences, as required for spatial imagination.
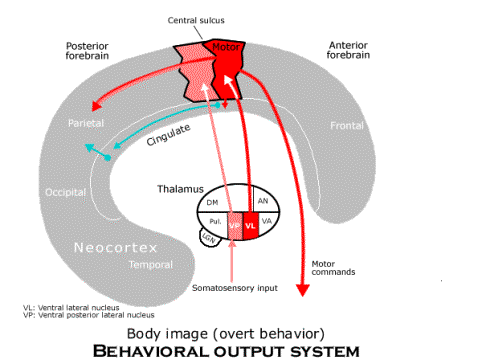
The body image for overt behavior includes the motor neocortex, supplementary motor neocortex and somatosensory neocortex, all of which are organized somatotopically, like an image of the body. The overt bodily image is the source of motor commands to all parts of the body, but it is mediated by a projection (not depicted) from the overt body image to the cerebellum and its projection back to the motor neocortex by way of a part of the ventral lateral nucleus of the thalamus. (The cerebellum evens out the gross motor commands of the premotor and supplementary motor areas both spatially and temporally so that the precise motor commands work together smoothly in generating the prescribed behavior in the body as a whole.) This overt body image located in the neocortex is also the source of the input about bodily condition that is used to construct the local image. Its projection (via association fibers) to the inferior parietal neocortex is used to combine telesensory images as the local image representing the spatial relations among object in the current local scene relative to the body. Finally, the overt body image also sends a projection to the anterior cingulate neocortex, and since it projects to the posterior cingulate, the overt body image provides the labels for recording memory images according to the locomotion and turning that are responsible for them.
When behavior is being generated overtly, both parts of the circuits through the corpus striatum are active. A behavioral schema in the frontal neocortex is used to set up behavior in the premotor area, and when the signal (from the centromedian nucleus of the thalamus) to the putamen calls for the behavior to be generated overtly, the schema is used to send motor commands to the body. (The centromedian nucleus is in a position to tell when to generate behavior overtly, because it is at the center of the thalamic nucleus receiving processes from all its nuclei as the thalamus synchronizes processing in all parts of the neocortex.) When behavior is overt, the anterior cingulate gyrus receives input from both the overt and covert body images, and thus, the labels attached to memory groups that are formed when behavior is overt can be used to recall those images in sequence as spatial imagination when behavior is covert.
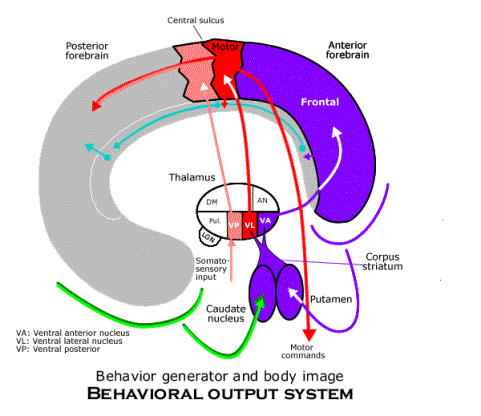
All the circuits combined are the subjective animal system of representation. The auditory system (in the superior temporal neocortex) and the tactile system (in the superior parietal neocortex) are not depicted.
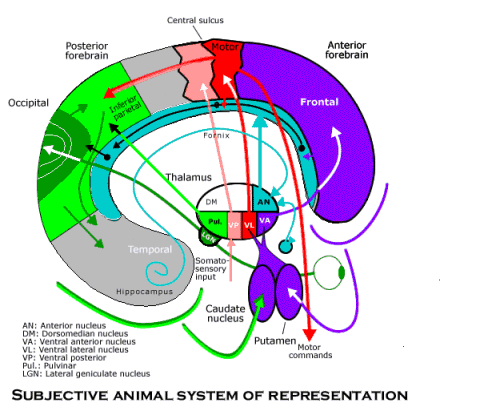
All that is needed to complete the subjective animal behavior guidance system is the goal selection system. It is also a complete circuit through both the thalamus and the neocortex. The route from the neocortex back to the thalamus so that a circuit can be completed is provided in this case by the amygdala. Located in the anterior temporal lobe, the amygdala receives the same information from throughout the neocortex that the memory circuit receives by way of the hippocampus, except that it passes through an ancient form of cortex surrounding it (the piriform cortex). Thus, the amygdala is in a position to attach desires to the objects (or read affective labels already attached to objects) that are used by behavior schemata to generate behavior in relation to them. The amygdala projects to the dorsomedian nucleus of the thalamus, which projects in turn to the whole frontal neocortex. A diffuse projection to most of the frontal area is all that is needed to activate appropriate behavioral schemata. A denser projection from the dorsomedian nucleus is received by the orbitofrontal region, and it sends association fibers (by way of the uncinate fasciculus) back to the piriform cortex surrounding the amygdala. Thus, the goal selection circuit is able to make sure that desires are attached to the right objects.
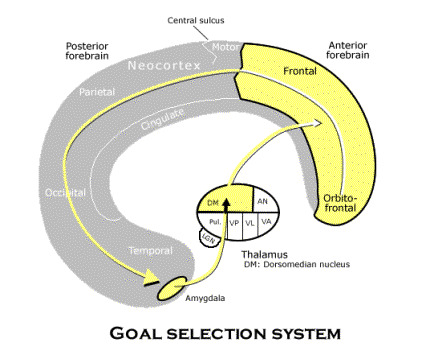
![]() Internalizing
the spatial structure of the world. Even telesensory
animals internalize a map of their territory. But their map plays very specific
roles in generating complex, “hardwired” instinctive routines in telesensory
animals. However, much of what is hardwired instinct in telesensory animals
is learned by subjective animals from the environment. That is partly what
makes the subjective level of neurological organization possible. As we have
seen, what makes learning so much more important in subjective animals is
a form of reproductive causation contained in individual brains with spatial
imagination by which behavioral schemata evolve by reinforcement selection.
(A similar mechanism has been described by Edelman,
1987, 1989, under the name of "neural Darwinism".)
Internalizing
the spatial structure of the world. Even telesensory
animals internalize a map of their territory. But their map plays very specific
roles in generating complex, “hardwired” instinctive routines in telesensory
animals. However, much of what is hardwired instinct in telesensory animals
is learned by subjective animals from the environment. That is partly what
makes the subjective level of neurological organization possible. As we have
seen, what makes learning so much more important in subjective animals is
a form of reproductive causation contained in individual brains with spatial
imagination by which behavioral schemata evolve by reinforcement selection.
(A similar mechanism has been described by Edelman,
1987, 1989, under the name of "neural Darwinism".)
The basic structure of the forebrain systems is set up in mammals by the biological behavior guidance system during embryological development, including the circuits of 2-D arrays that constitute the basic subsystems of the subjective animal behavior guidance system. But more neurons are created than are needed, and many neurons can make synapses more or less randomly with certain other neurons. The basic circuits line up 2-D arrays so that sensory input is registered in a way that can be used in generating behavior, and correspondingly, the mechanisms of the behavior generator are set up to make use of such sensory input in guiding behavior. But the specific neural connections needed to analyze and store sensory input in the most useful ways are lacking, and genetically prescribe behavior in response to sensory input is little more than reflexes built into in the behavioral output system and more or less random combinations of them.
Behavioral schemata in the behavioral output system evolve by a contained form of reproductive causation. As random variations on behavioral schemata are tried out, there is a natural selection of those that are successful in some way by a reinforcement that works through the memory circuit.
Though the memory circuit is depicted in the functional diagram as the basic structure of the sensory input system, it is realized in the brain in such a way that it is able to work the same way on neurons throughout the neocortex, including the behavioral schemata and body image in the anterior as well as the local image in the posterior neocortex. When some set of neurons is successful in some way, the memory circuit ties those neurons together in a memory group. That is, it instructs them to form synapses with one another so that if any crucial group of those neurons fire, the whole group will fire together. And with their synapses with one another strengthened, the whole group is more likely to be included as parts of other random variations on behavioral schemata.
Let us call such groups of neurons, which may be in the same 2-D array or in many different 2-D arrays, “memory groups.” Though the formation of memory groups depends on the memory circuit, once they are formed, they can be called up without any help from the memory circuit. The newly formed synapses among the neurons will make them act as a group when aroused by other neurons in the neocortex.
This increase in their likelihood of firing together again as a group is a form of natural selection of the group. It call it “reinforcement selection,” because the criterion for forming memory groups is, in mammals, the kind of desire satisfaction behaviorists were actually referring to in talking about reinforcement. The memory circuit is an ancient structure that includes the seat of desire, sometimes called the “limbic system,” and thus, success in satisfying desire in some way or other can easily be what triggers the formation of memory groups. In the case of learning behavioral schemata, success can easily be detected, for as we shall see, it is just success in internalizing some aspect of the world.
Mechanisms for identifying objects from the analysis of sensory input evolve in a similar way in the sensory input system. At the beginning of this phase of neurological development, groups of neurons respond to sensory input more randomly, and the memory circuit internalizes detailed structures that enable the sensory input system to recognize objects in sensory input and discriminate among them.
Telesensory input in any modality, such as vision or hearing, is registered in many different 2-D arrays. Those arrays may be determined to respond to them in different ways by the biological behavior guidance system, but the memory circuit, using built in criteria for success, can tie neurons together that are regularly active in processing sensory input in one way in one 2-D array with other neurons that are regularly involved in processing the same input in another way in another 2-D array. Thereafter, when sensory input is recognized in one 2-D array, such association fibers make it more likely that the input will also be recognized in the other 2-D arrays. Thus, processing in the various telesensory images work together to disambiguate sensory input.
At a later stage of neurological development, when mechanisms for recognizing objects of certain kinds have been established, this mechanism can also be used to identify particular objects. Different objects of similar kinds can be distinguished by tying a general memory group together with additional neurons in various 2-D arrays that are distinctive of it in a new memory group.
When new sensory input causes patterns of neuronal firings in many different 2-D arrays of the sensory input system that conform to such a memory group, the memory group is activated, and the sensory input system settles into equilibrium. (Since these areas of neocortex are all coordinated by the thalamus, such a settling into equilibrium may be what causes the distinctive “gamma oscillations” that are detected by EKGs when objects are recognized.)
For our purposes, what is most relevant about this contained form of reproductive causation is how the evolution of behavioral schemata internalizes the spatial structure of the world as the structural cause of spatial imagination. Though the behavioral schemata are certain groups of neurons in the frontal neocortex that the behavior generator uses to issue motor commands from the body image, what evolves are neural connections throughout the neocortex, because motor commands must be generated in relation to sensory input. These groups of neurons are the “organisms” (or primary structures) that evolve, because they are what go through reproductive cycles and have both essential structural effects. Random variations on them are naturally selected by success in reproduction, though in this case reproduction occurs when they generate overt behavior and it is reinforced through the memory circuit.
The basic schema has the part-whole complexity required of an organism that can evolve, since it is made up of motor commands to different parts of the body as well as different sequences of such sets of motor commands in time. Thus, many varieties of the basic schema are possible.
These schemata also do both kind of work that are required for organisms to evolve. Their non-reproductive work is the behavior they generate in relation to sensory input when they are firing. And they reproduce when the groups of neurons involved are signaled to strengthen and extend their synapses, that is, are reinforced for working together in the various circuits of 2-D arrays in a way that satisfies desire.
Thus, behavioral schemata evolve gradually, starting off simple, uniform and weak, and becoming complex, diverse and powerful. The ecological niches to which they adapt are all the various situations in which some special kind of behavior in response to sensory input is needed to satisfy a desire, that is, are reinforced.
Memory groups for identifying the telesensory images for situations in which behavior is reinforced are part of the selected behavioral schemata, that is, like discriminative stimuli. But since it is a faculty of spatial imagination, connections are also established between telesensory images and behavior that work the other way, so that when the behavior is generated covertly, telesensory images representing its effects are called up from memory.
In order to construct even a local image, the telesensory input must be registered in a way that indicates the bodily condition at the time, such as the direction the head and eyes are pointing when visual input is registered. That is a regular relationship in any given local scene viewed from a certain location, and thus, the memory circuit can establish their neural connections as a memory group. But in order for them to serve as a local image that can be recalled from memory, it must be possible for the behavior generator to use the combination of telesensory images recorded in memory as a local image to anticipate what telesensory images will occur when it behaves in various way, and that is a result of the evolution of behavioral schemata in the behavior generator.
Starting with whatever behavioral schemata are built into the behavior generator genetically, such as standing up, walking, focusing eyes together on an object, behavioral schemata for more complex or more precisely controlled behavior can be acquired by reinforcement selection of random variations on them. But since telesensory images are recorded in the local image according to the bodily condition at the time, schemata for generating those bodily conditions become labels for calling up those telesensory images in imagination. Thus, as behavioral schemata evolve, the subjective animal learns the motor commands to use in moving its head and body in order to make an object that appears to the right visual field move to the center of the visual field.
Though local images vary with the local scenes they represent, the same behavioral schemata for calling up particular telesensory images from them will work for them all, because bodily movements have similar effects in all situations. The behavioral schema that evolves for that function becomes, in effect, the structure for recording all local images, and the covert behavior for generating bodily conditions become labels for calling up the telesensory images in different directions. Thus, the evolution of behavioral schemata internalizes an aspect of the spatial structure of the world.
Once the ability to construct and recall local images has been acquired, much the same kind of process will internalize the structural causes needed to link them in sequences as a map of its territory (assuming that the cingulate gyrus contains a mechanism that can link memory groups in sequences). Given that behavioral schemata for locomotion and turning in relations to perception have been acquired, the key to spatial imagination is recording and recalling entire local images in a way that corresponds to such behavior.
Starting from a certain local scene in the territory, the effects of locomotion and turning on the local images encountered are determined by the spatial structure of the actual world. Thus, assuming that the memory for sequences of local images (in the cingulate gyrus) receives input about its overt locomotion and turning, its neural connections to the resulting changes in local images become labels for recording and recalling local images as a map of the territory. Using those behavioral schemata to generate covert locomotion and turning in relation to a given local image would, therefore, enable the subjective animal to explore the map of its territory.
Once again, however, the same kinds of behavior, turning and locomotion, work much the same way in any local image of the territory. Thus, as these behavioral schemata evolve, the memory for sequencing local images (and for shifting from one sequence to another when the body turns in a different direction) acquires labels that are used for every local image. That is a general understanding of the structure of space in terms of the effects of motion, and it informs even the local image, that is, enables the subjective animal to see its telesensory input from objects in the local scene as spatial relations that can change by motion. For example, as it learns to register local images according to turning, it learns that turning far enough brings it back to the same direction, and it can come to see how it would have that effect even in the local image.
The capacity to acquire structural causes from the environment eases the burden on the biological behavior guidance system to provide structural causes, but once the basic structure of the mammal’s higher level of neurological organization had been selected, evolution by reproductive causation would provide the additional structures needed to make this learning mechanism more efficient and complete.
The representation of the local scene is, in effect, a representation with polar coordinates, whereas the memory map of all the salient objects in the territory uses, in effect, Cartesian coordinates. This translation between polar and Cartesian coordinates is mathematically too complex to be provided genetically, but it can be acquired from experience. As the mammal experiences the consequences of moving in different directions from the local scene and finds the same objects showing up as a result of different combinations of bodily movements, behavior schemata for turning and moving in each direction evolve, so that later they can be used by the behavior generator as labels (in the anterior cingulate) to switch from one sequence of local images to another in exploring its subjective map. The mammal discovers, for example, the direction in which the body winds up moving as a result of various combinations of bodily movements, such as turning, backing up, rolling over, and the like, and so those combinations become packaged as behavioral schemata, or possible intentions, to move in certain ways in local scenes. They provide the translation from the "polar coordinates" of the local image to the Cartesian labels of the memory map. Only when a young mammal has learned the effects of turning and locomotion in the local scene is it able to start using the memory circuit to construct its own 3 .D memory map of the territory.
![]()