![]()
 6
Subjective stage. Subjective animals are multicellular animals with a higher level of neurological
organization than telesensory animals. They are all mammals. We will see why
mammals are inevitable and trace their gradual evolution. But as a result
of explaining the mechanisms built into their higher level brains, we will
also understand why there is a unity to the consciousness of mammals like
us, finally repaying in full one of the four mortgages we took out to use
spatiomaterialism as the ontological foundation for this argument.
6
Subjective stage. Subjective animals are multicellular animals with a higher level of neurological
organization than telesensory animals. They are all mammals. We will see why
mammals are inevitable and trace their gradual evolution. But as a result
of explaining the mechanisms built into their higher level brains, we will
also understand why there is a unity to the consciousness of mammals like
us, finally repaying in full one of the four mortgages we took out to use
spatiomaterialism as the ontological foundation for this argument.
The inevitability of an evolutionary stage following the evolution of telesensory animals depends, according to this ontological explanation of revolutionary change in evolution, on a higher level of neurological organization being both possible and functional. We have found good reason to doubt that such a higher level of neurological organization is possible in invertebrates. Their kind of embryological development is not able to try out the radical random variation required to organize neurons on a higher level of part-whole complexity. Or to put it positively, the main reason that such a radical random variation is possible in vertebrates is that their kind of embryological development is able to determine the fates of entire 2-D arrays of neurons as ordered units. That makes it relatively easy to organize such systems of neurons as parts of larger, complete circuits in the process of embryological development.
The mechanism of embryological development is, however, only part of the explanation of the possibility of mammals. Its possibility also depends on the way in which the more detailed structure of brains with the subjective level of neurological organization can be acquired from the structure of the world, that is, by learning from experience. Such learning is, as we shall see, a later phase in the process of neurological development in subjective animals, a stage during which detailed nervous structures evolve within the individual brain through natural selection by reinforcement (that is, as a form of evolution by reproductive causation that occurs within the mammalian brain).
If a further reason is needed for believing that the subjective level of neurological organization is possible, it will be found in following the actual evolution of nervous structures in the mammalian brain from the structures that were identified in the vertebrate brain in our discussion of the previous stage, stage 5. That is another way of proving that the structures to be identified in the mammalian brain actually have the functions that I will ascribe to them.
Both of these further reasons for believing in the possibility of the subjective level of neurological organization depend, however, on its function, that is, on how it opens up an entire, new range of powers that can evolve for controlling relevant conditions. Though I have already said that its function is to give animals spatial imagination, the time has come to explain what is meant by such a faculty of imagination and to describe functionally the systems and subsystems of the animal behavior guidance system that would give animals such a cognitive power. That is where we begin.
![]() Function
of the subjective level of neurological organization. Since
animal behavior acts on other objects in space, its power can be increased,
as we have seen, by adapting the behavior to spatial aspects of the situation
in which the animal must act. That makes animals better able to control the
conditions affecting their reproduction. One of those aspects is the way that
the structure of the space, by helping to constitute what happens in the region,
imposes regularities on how such objects can change location and interact
with one another, or what I have called spatial causation (including
how it helps cause local regularities ontologically as well as global). That
aspect is, perhaps, better described as a spatio-temporal aspect of the situation,
since it includes regularities about change. In any case, taking advantage
of it in guiding their behavior to act on other objects in space is the basic
function of spatial imagination. The function of the subjective neurological
level is, in short, to understand spatial causation.
Function
of the subjective level of neurological organization. Since
animal behavior acts on other objects in space, its power can be increased,
as we have seen, by adapting the behavior to spatial aspects of the situation
in which the animal must act. That makes animals better able to control the
conditions affecting their reproduction. One of those aspects is the way that
the structure of the space, by helping to constitute what happens in the region,
imposes regularities on how such objects can change location and interact
with one another, or what I have called spatial causation (including
how it helps cause local regularities ontologically as well as global). That
aspect is, perhaps, better described as a spatio-temporal aspect of the situation,
since it includes regularities about change. In any case, taking advantage
of it in guiding their behavior to act on other objects in space is the basic
function of spatial imagination. The function of the subjective neurological
level is, in short, to understand spatial causation.
![]() Spatial
imagination. In order to show that such a faculty of spatial imagination
would make animals inherently more powerful than telesensory animals, let
us recall the cognitive powers of telesensory animals. Not only do telesensory
animals have the capacity to guide behavior in relation to objects that can
currently be perceived, but they also have the capacity to construct a map
of the territory by which they can guide behavior in relation to objects that
are not currently perceived.
Spatial
imagination. In order to show that such a faculty of spatial imagination
would make animals inherently more powerful than telesensory animals, let
us recall the cognitive powers of telesensory animals. Not only do telesensory
animals have the capacity to guide behavior in relation to objects that can
currently be perceived, but they also have the capacity to construct a map
of the territory by which they can guide behavior in relation to objects that
are not currently perceived.
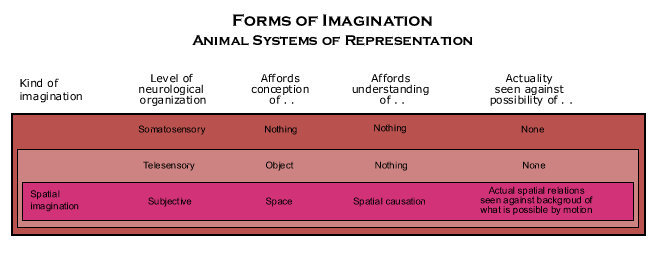
The territorial map used by telesensory animals, such as bees and birds, is basically, however, just an elaboration of the complex instincts that have evolved in relation to the telesensory representations for detecting of kinds of objects in order to choose which of incompatible kinds of behavior to generate toward them.
The nervous mechanisms set up during embryological development is a structural cause not only for using input from the bodily condition to register telesensory input in a way that reveals the locations of perceivable objects in space, but also for the instinctive routines that use telesensory input to construct and use a map of its territory. The telesensory map is little more than a set of computational tricks that take advantage of the geometrical structure among salient objects on the surface of the earth to revisit energy sources and return to a nest or home base. Though computations on quantities like distance and azimuth and one-dimensional, stimulus-response chains for retracing pathways are effective, the isomorphism between states of the brain and the basic three dimensional structure of space is only implicit in them. Nor does it represent the effects of locomotion on the relations of objects in the territory, but merely guides actual locomotion.
These fixed action patterns built into the structure of the brain are called "programs of behavior" by J. Z. Young (1978, and 1981, p. 362), though Young assumes that they explain the behavior of mammals as well as that of other vertebrates and invertebrates.
The structure of space is, however, so important in determining the outcome of interacting with other objects in space that animals would be even more powerful, if the animal system of representation took greater advantage of it. Since material objects are substances that coincide with substantival space, they can change location, as we have seen, only by moving across space as time passes. And since space has a three dimensional structure, the effects of motion on the relations of objects in space is determined by the object’s speed and direction relative to them in space. This is a spatio-temporal aspect of the world, and adapting animal behavior to it would make animals more powerful.
Telesensory animals already adapt the spatial aspect of animal behavior to spatial aspects of the world. That is, they use their animal behavior over time to impose a kind of geometrical structure on the thermodynamic flow in a region that would otherwise require a giant machine. But instead of the unchanging geometrical structure of a huge material object, the structural cause is just an animal with a nervous system to guide its behavior so that it is in the right place at the right time to make things happen in the region that otherwise would not happen. Thus, it is an active, energy consuming way of imposing a geometrical structure that causes irreversible changes in the region that would not otherwise occur. This telesensory adaptation of the spatial aspect of animal behavior to spatial aspects of the situation is already to use a spatio-temporal aspect of behavior to control what happens to material objects located in the region.
Though animal behavior as such has a temporal aspect as well as a spatial aspect, telesensory animals do not adapt the spatio-temporal aspect of their behavior to the spatio-temporal aspect of the world. They adapt it only to the spatial aspect of the world. When they act on the perception of the object, it is the perception of the object in its current location. And if the other object is moving, that means guiding locomotion in relation to its current, albeit changing, position. However, when they act on their map of the territory, they assume that the locations of objects in their territory do not change. Their map gives them power over other objects by guiding locomotion in relation to unperceived objects whose locations do not change.
Animals would be more powerful, however, if they could adapt the spatio-temporal aspect of their behavior to the spatio-temporal aspects of the world.
If they could anticipate the consequences of the motion of objects on their relations to other objects in space, they could guide locomotion in relation not only to distant, unseen objects, but also in relation the future locations of objects that can move around in space. In chasing another animal, for example, they could move in the direction of its future location, rather than where it is currently perceived.
The capacity to anticipate the consequences of its own locomotion on the relations of their bodies to other objects in space would enable them, for example, to figure out an alternate route to its destination when the usual path is blocked, whereas the telesensory animal would be left with rigid routines that simply no longer work.
These examples emphasize the way in which anticipating the consequences of motion gives the animal foresight. But anticipating the consequences of motion is more generally useful, because, as we shall see, it is an understanding of the structure of space which makes the animal better able to take advantage of purely spatial aspects of the relations among objects in its territory than telesensory animals. In chasing another animal, for example, it could not only project its future location from its current motion, but also figure out where the animal might have gone in the territory when it can no longer be seen.
"Subjective" is an appropriate name for animals like this, because spatial imagination depends on internalizing the spatial structure of the world in a way that gives them an understanding of the structure of space. It depends, as we shall see, on a faculty of imagination in which sequences of memory images represent the consequences of locomotion relative to certain starting points. That makes it possible to think about spatial relations in terms of the consequences of locomotion (and motion generally), that is, as a spatio-temporal aspect of the world. That is to understand intuitively the role of space as an ontological cause of regularities about change, or “spatial causation.” (The use of sequences of image over time to represent what is possible means that this faculty might better be called “spatio-temporal imagination.” But it means that spatial imagination also includes the ability to record in memory and call up in imagination other spatio-temporal aspects of the world, such as regularities about change that do not have to do with motion, including how other animals behave).
Mammals have, therefore, a conception of space. They understand intuitively that space has three dimensions and what that means about the possible spatial relations among objects in space. They see other objects as being located in space. They even see their own bodies as having a location elsewhere in the same space as the other objects they perceive in space.
The subjective animal system of representation gives animals, therefore, the conception of the object as being located in space. But since location in space means having particular spatial relations to every other object in the world, spatial imagination is not merely a representation of the object as in space, but also a representation of the whole world, at least, in principle. And it is a world in which it can see its own body as just another object located in relation to every other object in the world.
This faculty of imagination enables animals to see the actual against the background of the possible, in this case, the actual locations of objects in space against the background of what is possible by motion. What space makes possible is presented as sequences of images along with perception (or the images derived from current sensory input), and thus, the possible has an appearance to the animal. The mammal’s animal representation of space is explicit.
None of this is even conceivable by the telesensory animal, because the location of the object is only implicit in its animal representation. When the telesensory animal focuses on the object, its location in space is not represented in the images caused by telesensory input. Its location is contained as a mere code for generating locomotion in relation to the object, should it choose to approach or avoid it, that is, as a disposition to move in certain ways. Thus, the telesensory animal is unaware that its own body is another object with a different location in space from the object being perceived. Indeed, it is not even aware that it has a body, except implicitly in special kinds of behavior, such as preening in birds, that has been selected for taking care of its own body. Otherwise its body is simply how it moves in relation to other objects and acts on them. It is as if the telesensory animal is its body, whereas the subjective animal is a subject to whom its body has an appearance, if only the appearance of being located in space relative to other objects.
Subjective animals require a higher level of part-whole complexity in the nervous system, because imagination depends on the capacity to generate behavior covertly and, by that covert behavior, to recall from memory the telesensory images that would result from those bodily movements. How the mammalian brain differs from the non-mammalian vertebrate brain is described below in detail, but first, the basic difference between the brain mechanisms of telesensory and subjective animals will be explained in functional terms.
Indeed, as an introduction to that functional description, the difference can be suggested by contrasting two kinds of computers.
Telesensory animals are comparable to the computer-guided robots found in factories and that have been used for exploring other planets. They can be programmed to generate routines in specific situations, but they rely on certain cues or landmarks to get around in space and, thus, can easily be confounded by changes in the locations of objects.
Subjective animals, by contrast, are comparable to computers that generate for beings like us what is called "virtual reality". In the computer memory are stored images of what would be seen if the human head were pointed in certain directions at each location for a whole set of locations to which a body can move, so that when sensors attached to a human body detect such bodily motion, they call up the appropriate image from the computer memory and project it onto a screen in front of the eyes, giving a human subject the appearance of moving around among the objects in some region of space. It generates an uncanny sense of being in an imaginary world, because the human being is playing the role of generator of "covert behavior" exploring a memory that is supplied by the computer, just as real covert behavior explores the memory of the subjective animal system of representation.
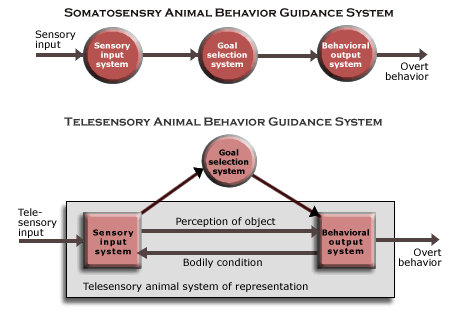
![]() Functional
diagram of subjective animal behavior guidance system. The various
functions and subfunctions of the subjective animal system of representation
are depicted in the accompanying diagram of the subjective animal behavior
guidance system. Though neural mechanisms serving the function of selecting
the kind of behavior are also essential to the animal behavior guidance
system, only the mechanisms serving the sensory input and behavioral output
subfunctions are involved in the animal system of representation (all the
parts with the gray background). The higher level of neurological organization
is suggested by the contrast with the diagrams of the somatosensory and telesensory
animal behavior guidance systems (which are also provided for comparison.)
Functional
diagram of subjective animal behavior guidance system. The various
functions and subfunctions of the subjective animal system of representation
are depicted in the accompanying diagram of the subjective animal behavior
guidance system. Though neural mechanisms serving the function of selecting
the kind of behavior are also essential to the animal behavior guidance
system, only the mechanisms serving the sensory input and behavioral output
subfunctions are involved in the animal system of representation (all the
parts with the gray background). The higher level of neurological organization
is suggested by the contrast with the diagrams of the somatosensory and telesensory
animal behavior guidance systems (which are also provided for comparison.)
In the subjective system, the sensory input and behavioral output systems both depend on an interaction of at least three brain mechanisms, each at the same level of neurological organization as the whole sensory input or behavioral output system of telesensory animals. Indeed, as the diagram suggests, each is a complete circuit of such mechanisms.
The brown squares and lines are basically the systems and causal connections serving the functions that are also served by the telesensory animal system of representation (including telesensory input, input about the current bodily condition, their combination as the perception of the object, and how that is responsible for overt behavior when some kind of behavior is selected). The higher level of part-whole complexity in the subjective sensory input and behavioral output systems makes it possible for them interact in the more complex way that constitutes imagination.
The red circles and lines (in each brown square) represent the systems and causal connections serving new functions required by spatial imagination (including the body image, covert locomotion, memory, the local image, the sequences of images, or spatio-temporal images, called up from memory by convert locomotion, and the behavior generator). Here is how it works as a faculty of spatial imagination.
(The three systems in each complete circuit include a thalamic nucleus, at least one region of neocortex, and a third structure of some kind, which is suggested by the two circles and a little ball. The neural mechanisms serving these functionally described systems are identified in the next section, on the structure of the mammalian brain.)
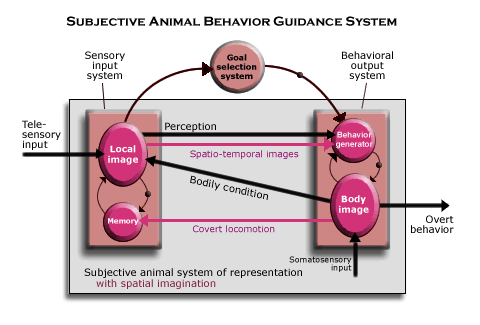
Local image. The telesensory input registered by the sensory input system includes all the telesensory modalities. Vision and hearing are most central, though olfaction is still used. Input about the current bodily condition has the same function with respect to telesensory input as it does in telesensory animals, except that by handling all the telesensory modalities together, it can construct a more complete representation of the current scene, or what is labeled as the “local image.”
Though the local image represents only objects that can be perceived from the animal’s current location, it also represents some objects that are not currently causing telesensory input. Telesensory input from objects located behind the animal at the moment, for example, is registered in short-term memory according to the bodily condition at the time it was perceived, while the objects on which the eyes focus or that are making noise are currently causing telesensory input. But since these telesensory images are all part of the local image, objects are perceived as being located relative to one another in the current scene, and behavioral output can be adjusted to them all (when some kind of behavior is selected).
Though the local image is a complex of many telesensory images which can guide behavior in relation to many different objects, it is the unit that becomes connected in sequences as the subjective animal’s territorial map in spatial imagination.
Body image. Overt behavior is caused by motor output to muscles throughout the body. The behavioral output system is labeled the “body image” because commands for overt behavior are sent from a representation of the body. The body image is a spatial structure in various 2-D arrays of neurons that represents somatotopically all the parts of the body whose behavior it controls, and it receives somatosensory input from all the same parts of the body, indicating the positions and motion of limbs, the stress of muscles, tactile sensations from the skin, and pain from injuries.
Tactile sensations reveal information about objects that contact the body, and since the stance of the body is represented, it reveals information about the location of the other object causing the somatosensory input. (Such information is registered as part of the local image by way of its input about the current bodily condition, though there are reflexes that can trigger behavior locally without involving the selection system, for example, in response to intense heat or pain.)
Behavior generator. Most motor commands in the body image are generated, however, by another part of the behavioral output system, labeled the “behavior generator.” The behavior generator contains schemata for all the various kinds of behavior that may be selected, and depending on its input from the goal selection system, the behavior generator uses one of them together with the perception of the object as located in space (that is, the local image, its input from the sensory input system) to generate precise motor commands that are adapted to the locations of relevant objects in the current scene. Thus, behavior may be guided in relation to perceivable objects even though they are not currently causing any telesensory input.
Moreover, the behavioral schemata used by the behavior generator may involve many steps, because the behavior generator has feedback from the motor commands issued by the body image by which each motor command triggers the next in a sequence, generating temporally complex patterns of behavior. There is a wide range of variations on behavior that may be included in such behavioral schemata, since motor output sends commands separately to each part of the body.
Thus far, the subjective animal system of representation serves basically the same functions as the telesensory animal with respect to currently perceivable objects, though in a more complete way, since its higher level of neurological organization integrates all the kinds of telesensory (and somatosensory) input as a local image on the input side and can generate a great variety of long sequences of complex motor commands on the output side. What is new is spatial imagination. Though spatial imagination serves the same function as the telesensory map of the territory animal, it is so much more complete that it represents an aspect of the world that inevitably escapes telesensory animals: regularities about how motion and locomotion affect the relations of objects in space. That is the source of the subjective animal’s conception of space.
Memory. The new subsystem in the subjective animal’s sensory input system is a special kind of “memory” which is able to record images in sequences so that they can be called up in the same sequence. The images may be just telesensory images of an object, for example, recording what happens as another animal behaves. But they can be entire local images. That is, various telesensory images representing a local scene (with each registered according to the bodily condition relevant to receiving it) can be recorded together as a memory unit, and local images can then be recorded in sequence according to the direction of its locomotion. The effects of locomotion in different directions from any given scene are recorded in different sequences.
Overt locomotor behavior is registered, not only in the local image by way of input about the current bodily condition, but also directly in the memory system by way of the “covert locomotion” connection. The direct connection to memory is what is used to label local images in constructing the map.
Covert locomotion. What calls up the local images recorded in this way is a new capacity of the subjective behavioral output system, the capacity to generate behavior covertly as well as overtly. In covert locomotion, the same motor commands that would otherwise cause locomotion in a certain direction relative to a local image are generated, but in a way that does not affect the body. It affects only the memory based in the sensory input system. Since local images have been recorded as units labeled by the appropriate bodily condition (mainly the stance of the body, head, eyes, ears and the like), and since local images have been connected in sequences according to the bodily condition at the time (mainly the direction of locomotion relative to the local scene), covert locomotion calls up the sequence of local images that would be encountered if the animal actually moved in that direction. The local images called up from memory are projected in a sequence over time to the behavior generator in the same way as images of perception, though they are just memory images (labeled “spatio-temporal images” in the diagram) and can occur faster. Thus, the behavior generator can use the local images from memory to generate covert locomotion in relation to unperceived objects in the same way as perceived objects, drawing on its schemata for various kinds of locomotion and generating motor commands in relation to objects in the imagined local scene (represented by the local image).
Together, such sequences of local images serve as a map of the animal’s territory. For example, if a local image that turns up in exploring one sequence by covert locomotion is registered as (or simply recognized as) part of another sequence of local images representing locomotion in another direction, the subjective animal can change the direction of its covert locomotion relative to that local image and anticipate what would happen from turning at that location in the other direction. In familiar territory, such sequences of local images are richly interconnected, but in any case, they give the subjective animal a map of the relations of objects in its territory. The subjective map is different from the telesensory map, because it can guide covert as well as overt locomotion in relation to goals picked out by the selection system. Thus, far from being merely a conditioned stimulus-response chain that leads the telesensory animal from one location to another as parts of a instinctive routine for controlling a specific relevant condition, memory is a potentially complete, general purpose map of all the objects in its territory which can be explored in imagination by covert behavior.
Evolution of behavioral schemata within the brain. Locomotion, covert or overt, is generated by behavioral schemata, and not only do subjective animals have many different ways of moving and turning in relation to other objects in space, but they also have other kinds of behavior. Though schemata for basic forms of behavior may be built into the structure of the brain (indeed, must be in the case of mammals that have to be capable of locomotion from birth), they can also be learned. And in any case, the consequences of acting on them is learned, and thus, how to adapt them to the situation is learned. Learning transforms the subjective animal’s map of its territory into an understanding of the structure of space, and we can see why, by recognizing that learning is a from of evolution by reproductive causation. Subjective animals learn as behavioral schemata evolve in the brain as the result of a form of natural selection that I will call “reinforcement selection.” Learning is a later phase of neurological development that occurs when the animal is interacting with the world, and its effect is to internalize certain aspects of the world in the skills and imagination of the subjective animal. Let me describe how learning can be seen as a contained form of evolution by reproductive causation, setting aside until later discussion of the neural mechanisms that serve these functions.
The “reproducing organisms” in this case are the behavioral schemata used by the behavior generator to generate motor commands in the body image. They go through reproductive cycles in the sense that, once they have generated behavior, they can do so again.
Variations on behavioral schemata are possible, because behavior is generated by motor commands to all parts of the body from the body image. Not only can highly varied (and finely tuned) kinds of behavior be generated, but such complex motor commands can also be generated in different sequences over time as parts of a single action. By rearranging the motor commands, variations on behavioral schemata can be generated more or less randomly.
The natural selection of behavioral schemata is made by their being recorded in memory. Such recordings increase the likelihood that the same kind of behavior will be generated again. (Basically, the synapses among the neurons used to generate the behavior are strengthened.) The behavioral schema reproduces when it is used again to generate behavior.
Since behavioral schemata are used to generate behavior in conjunction with the perception of the object acted on, they are recorded together with telesensory images of those objects in the local image. Two kinds of connections are established between telesensory images and behavioral schemata. Telesensory images of objects (or their local images) that precede the behavior may help trigger the behavioral schemata, when they recur in perception. And when behavioral schemata are used to generate covert behavior, they call up telesensory images (or local images) of what results. These connections are the way local images are recorded in sequences, so that they can be called up as spatial imagination, by locomotion and turning.
The criterion for natural selection (that is, recording it in memory) is basically whether the behavior generated by the schemata satisfies a relevant desire. The recording is made by the memory circuit, and its criterion comes from its relationship to the goal selection system. Random variations that do not succeed in satisfying desire are not recorded.
In learning basic kinds of behavior, the relevant desire is mainly just that its expectations about telesensory images be confirmed by what shows up in perception. The ecological niches are basic behaviors required to perceive objects in different directions and to move from one local scene to another, and the behavioral schemata that are adapted to those ecological niches internalize some basic aspect of the world.
This contained form of reproductive causation is the mechanism, in mammals, that behaviorists, like B. F. Skinner, called “operant conditioning.” What they called “operant behavior” is behavior generated by behavioral schemata. What they called “reinforcement” is natural selection by recording the behavioral schemata in memory. What they called “discriminative stimuli” are the telesensory images (or entire local images) that help trigger the behavior again.
Explaining operant conditioning in terms of the subsystems of the subjective animal behavior guidance system enables us to see this important kind of learning as a form of gradual evolution. Behavioral schemata start off simple, uniform and weak, and as a result of “reinforcement selection,” they become complex, diverse and powerful in generating behavior that can satisfy desire. The gradual evolution of behavioral schemata is the increasing power of the animal to control relevant conditions.
Though it is contained within the individual brain, it is a form of reproductive causation. Not only is natural selection made by success in reproduction, but reproduction is also what forces a selection to be made. The ecological niches in which behavior schemata can complete their reproductive cycles are situations in which the animal must behave in some way, and the reproduction of one behavioral schema (by its reinforcement) in such an ecological niche tends to cause a “scarcity” of opportunities for other behavioral schemata to be generated, because the animal can behave in only one way at a time as situations of that kind arise. To be reinforced, a behavior schema must generate its behavior at some time before its memory group fades away from disuse, and thus, the scarcity is caused by how reproductive cycles add up in time (rather than by how reproductive cycles add up in space as time passes, as in biological evolution). But as random variations on behavioral schemata are tried out in such an ecological niche, those that satisfy the desire most efficiently are reinforced, and thus, “reinforcement selection” causes a stage of gradual evolution in which each behavioral schema becomes maximally efficient at satisfying some desire in certain situations (its ecological niche), and the combination of behavioral schemata at the ecological level, or the animal’s repertoire of behavior, comes to satisfy all its desires in all the situations it encounters as fully as possible with the least effort required. At both the individual and ecological levels, therefore, behavioral schemata change in the direction of natural perfection, that is, doing the most with the least.
What the doctrine of operant conditioning leaves out, however, is the faculty imagination. Though operant conditioning may be adequate to explain learning in telesensory animals, it leaves out important aspects of learning in subjective animals that depend on spatial imagination.
The relevant consequences of behavior are not limited to whether the behavior is reinforced or not. When a behavioral schema is reinforced, what happens, that is, the telesensory images (or local images) of what happens, are also recorded in conjunction with the behavioral schemata. Thus, when a behavioral schema is generated covertly, those images are called up in memory. Thus, when the subjective animal moves or turns covertly in the local scene, it can anticipate the objects and other local scenes it will encounter. This is a map of the animal’s territory, because when covert locomotion or turning is made relative to a local image, it calls up other local images relative to it. Thus, the subjective animal can explore its territory in imagination.
But the capacity to record and recall local images according to locomotion is not just a map of a particular territory. Learning to construct and use memory maps gives the subjective animal a general conception of space by which it can understand any relations in space by imagining the kinds of motion that would be involved in traversing or changing them. This general understanding comes from how the memory system reinforces behavioral schemata. The same kinds of locomotion have the same kinds of effects in similar local scenes. For example, when confronted with a large object, moving around it to the right and then turning left will give one a different view of the object. Or when one encounters a hill, climbing it will give one a view of what is on the other side. Two right turns in succession sends one in the opposite direction. Thus, by recording local images with similar structures together as the same generalized local image, covert locomotion becomes a general understanding of the structure of space, because the subjective animal has a “map” of the consequences of locomotion that would work for all local scenes of that kind. And by generating a sequence of locomotor schemata in sequence, it could anticipate the general consequences of combining them in certain ways.
In other words, the spatial structure of the world is internalized in spatial imagination because of how the memory system records behavioral schemata. It is the spatial structure of the world that accounts for the regular connection between kinds of locomotion or turning and the kinds of changes that occur in local images, and a behavioral schemata evolves by reinforcement selection, the connections in the memory system internalize those general relations. Their internalization is the subjective animal’s conception of space.
Understanding the structure of space. Though spatial imagination is based on a map of the territory in which the consequences of locomotion are represented by sequences of local images, the way that behavioral schemata evolve within the brain by reinforcement selection means that it is also a more general understanding of the structure of space. That is, it internalizes not only the particular relations among material objects in the animal’s territory, but it also internalizes the structure of space itself. In both cases, it is spatial imagination, because the understanding comes from the sequences of local images that can be called up covert locomotion (and turning).
What would happen because of locomotion (or the motion of any object) relative to objects in a local scene can be anticipated, because spatial imagination, that is, the capacity to generate behavior covertly, calls up from memory sequences of local images that represent the consequences of such behavior. The local images can be particular or general. Covert behavior acts on memory in similar ways in both cases. The difference is whether covert behavior engages with the detailed, particular local images that are the map of its territory or it operates on more generalized local images, which contain only certain relevant features. (They are actually groups of more specific memory images. We shall see how such general memories are possible when we take up the nervous mechanisms that serve these functions.)
Since spatial imagination gives the subjective animal an understanding of the effects of locomotion and turning in any space, it can see the objects in any local scene as being located in space. That is, the way that general local images are called up in sequences up by covert locomotion represents general regularities that are caused ontologically by the structure of space, and since those regularities hold not only among local scenes, but also among the objects within any local scene, the subjective animal has an intuitive understanding of the constraint that space imposes on change. That is the sense in which it perceives the object as being located in space. It is a conception of the structure of space.
Since spatial imagination gives the subjective animal a conception of space, the subjective animal sees not only other objects, but also its own body, as located in space. The local image is organized relative to the body, since telesensory images are combined as parts of it according to input about the current condition of the body. But since the subjective animal eventually acquires an understanding of the structure of space, that understanding applies to objects within the local image, and that includes the ability to think about the body as another object in space. The subjective animal can imagine not only its own body moving relative to other objects in the local scene, but also the effects of other objects moving in relation to its own body — as well as relative other objects in the local scene. Thus, even the body itself is represented as an object in the local image.
To sum up, we might even say that the subjective animal is able to see the world as a whole. To be sure, its map of the relations of objects in space is limited to objects in its territory. But it has a way of understanding spatial relations that can be extended, in principle, to include every object in the world. Thus, given how the world has a wholeness that comes from the structure of space, the mammal sees the world as a whole. Since such subjective animals have an intuitive (nonlinguistic) understanding of how a world of objects in space is inherently whole, they can perceive objects as part of a single whole.
Behavioral evidence of spatial imagination. As suggested at the outset, spatial imagination makes subjective animals more powerful than telesensory animals in various ways.
Spatial imagination give the subjective animal a map that not only guides instinctive behavior in relation to objects in its territory, but also enables the animal to see particular objects as having particular locations relative to others. It is a set of beliefs about the whole world that is based on being able to understand the consequences of motion relative to them. And since it is a general purpose map of the animal’s territory, rather than just an elaboration of instinctive routines for acquiring energy, mating, avoiding predators or the like, it can be used in pursuit of goals that were not relevant to its being acquired.
It enables subjective animals to plan new routes to goals when their paths are blocked, to figure out how to escape predators, to predict which direction prey might run, to choose the best way to catch them, and the like. It is the "inner world" to which Dennett (1975) was pointing in his famous essay, "Why the Law of Effect Will Not Go Away", when he said (paraphrasing Popper) that animals have an inner world that allows "hypotheses to die in their stead".
It gives subjective animals a primitive way of reasoning, for they can draw inferences about a world of objects in space. The animal system of representation is based on the assumption that objects do not come into existence or go out of existence, and with spatial imagination, that assumption can, for example, be used to infer where an object that it is seeking is located by knowing where it is not located and has not been. As a form of understanding, therefore, spatial imagination is a way of using current beliefs to form new beliefs, that is, a way of making inferences.
There is empirical evidence that spatial imagination makes mammals more powerful than non-mammalian vertebrates.
But it should be noted that not everyone agrees that mammals are more intelligent than non-mammals. In a study by Macphail (1982), the behavioral criteria used to argue that mammals are not measurably more intelligent than non-mammalian vertebrates do not take into account the capacity to get around in space. And although Gallistel (1990) recognizes that memory files are labeled spatially, he does not recognize the difference between the instinctive computational routines used by telesensory animals and a subjective memory based on the three dimensional structure of space. In general, there is no recognition in the literature of mammalian superiority.
Experiments demonstrating that rats use a representation of the spatial relations of objects in the local scene to orient their locomotion are reviewed by Gallistel (1990, Ch. 6). And rats that were not merely conditioned to the unusual environments that Gallistel discusses, but raised in them, should make even better use of the cues about spatial relations of objects in the region, because they would have internalized a map using input about the kinds of bodily conditions required. In any case, it is easy for mammals to learn mazes, because the mechanisms of spatial imagination acquired from experience with a world of objects in space would enable them to construct a new map for the maze.
There is evidence of inferences based on spatial imagination in how mammals in familiar mazes take advantage of apparent new shortcuts to the goal in otherwise familiar mazes. For example, if, at some point in a maze that a rat has learned, the normal route is blocked and barriers are removed both in the direction of the goal and in the opposite direction, the rat will take the new route to the goal, not the other one. (See Charles Taylor, 1964, pp. 161.196. These experiments pose a problem for traditional learning theory, which explains maze behavior as chains of discriminative responses. See also Gallistel 1990, Ch. 5.)
Even birds do not take advantage of shortcuts (when walking) through mazes, despite being able to learn mazes almost as quickly as mammals. In fact, once birds have learned a maze, it is surprisingly difficult for them to learn changes in it at all. Although birds have a mammal-like memory circuit, this suggests that, at least, when it comes to walking, it is not a spatial imagination that gives them an understanding of the structure of three dimensional space.
Subjective animal behavior guidance system. The faculty of spatial imagination serves as the animal system of representation in subjective animals, and in order to be of use in guiding behavior, the right kind of behavior for each situation must also be selected. Indeed, selecting between incompatible kinds of behavior is the original function of the animal behavior guidance system. And imagination also makes subjective animals better able to tell which goal to pursue in the situation.
The goal selection system. The kind of behavior generated must be made to depend on the situation, either the kinds of objects that are present in the local scene or the current needs of the body (or both). In subjective animals, goal selection is not just a matter of instinctive routines, but depends on desires, which are dispositions to behave in certain ways. Thus, its goal selection system might also be called the “affective system.”
The seat of desire is the goal selection system, for it registers of the various needs of the body and keeps track of the success of behavior in satisfying each of its desires.
Special input from the body is supplied to the goal selection system by the biological behavior guidance system (not represented in the functional diagram).That keeps the goal selection system informed of the body's need for food, water, sleep, mating and the like.
Objects represented in the sensory input system also have affective labels, which can arouse desires in the goal selection system or determine which object to pursue when a desire is already aroused. Some affective labels are instinctive, that is, built into the structure of the animal behavior guidance system, such those for avoiding certain predators and mating. Other affective labels are attached to objects by the goal selection system according to past experience. That is, the goal selection system can use the results of behavior to label objects by their capacity to satisfy desire, so that objects can be recognized for their capacity to satisfy desire when they are encountered again.
In either case, when the object turns up in the sensory input system, the desire may be triggered. But if the desire is aroused by bodily needs, telesensory images of the relevant objects may be called up from memory by their labels.
Whether the desire is aroused by sensory input or bodily needs, the goal direction system selects the appropriate kind of behavior by strengthening a certain behavioral schema in the behavior generator. (This is represented in the functional diagram by the projection from the goal selection system to the behavioral output system.) In other words, in terms of behavior, desires are dispositions to behave in some way, such as eating, mating, fighting, fleeing, and sleeping. But since animal behavior acts on objects in space, the desires that have this effect on the behavior generator must also be attached to an object.
When the object represented by sensory input is what arouses the desire, the object for the behavior schema to act on is already determined. If there is an obstacle in its way, it poses a problem that the subjective animal may be able to solve by spatial imagination. For example, it may use covert locomotion in its map of the territory to figure out a route that would put its body in the right location to act on the object as it is disposed to.
If the desire is aroused by bodily needs, the behavioral schemata it calls up may include searching for objects that will satisfy it, as in foraging for food and finding shelter. Or it may call up images of objects that will satisfy it, and if such images happen to be part of some local image in the map of its territory, the mammal can explore its map covertly to set up the locomotion that will lead to it.
In either case, instead of the telesensory animal’s rigid instinctive routines, desires set practical problems to be solved by using spatial imagination to imagine alternatives, and so subjective animals would be able to behave in a more flexible and adaptive way when necessary.
The goal selection system is the third essential subsystem of the animal behavior guidance system (along with the sensory input and behavioral output systems), and though it is not depicted in the functional diagram, it is also on the subjective level of neurological organization, like the other systems. There is a seat of desire, and it interacts through a complete circuit of connections with the other two subsystems. That is how it is able to attach the desires, or dispositions, to the right objects.
Sleep and dreaming. Dreaming, or REM (rapid eye movement) sleep, evolves with mammals (although there is some evidence of it in birds), but it has long been puzzling what its function could be. Given the evolution of spatial imagination in subjective animals, however, its function could be to try out random variations on behavioral schemata so that reinforcement selection can be more selective in attaching the right desires to the right objects in the right situations.
The attachment of desires to objects is a crucial function in behavior guidance, because it sets the goals for animal behavior. The animal will not be as powerful as possible, for example, if it fails to attach the desire to eat to objects from which it can acquire food, and it can be disastrous to attach the desire to eat to a predator. Furthermore, the task becomes increasingly complicated as the behavioral schemata for satisfying certain goals become increasingly complex. In the behavior schemata for attaining certain goals, behavior towards several objects of various kinds may be involved. Special routes through the territory may be needed to acquire food in certain ways. Some locations or objects may be dangerous. And social relations require different attitudes toward different members of the group not only in the family but also when there is a dominance hierarchy.
Reinforcement from satisfying desire is responsible both for attaching desires to objects and for strengthening behavioral schemata for satisfying them. Indeed, they are the same thing, because behavioral schemata that are naturally selected by success in satisfying certain desires include images of the objects and situations in relation to which the behavior generator sets up the motor commands. But natural selection by reinforcement is not very selective when success comes from subtle variations on complex behavioral schemata or accidents. Established behavioral schemata may be more powerful with their parts rearranged. Thus, reinforcement selection would be more effective in making behavior maximally powerful, if it had a wider range of random variations from which to choose, as long as it did not risking making disastrous mistakes.
Spatial imagination makes it possible for subjective animals to try out random variations on behavioral schemata in a safe way. Indeed, the regular use of spatial imagination in that way may be the function of dreaming.
Behavioral schemata are connected with both the desires and situations in which they are acted on, but they are often complex behavioral schemata, and thus, if the subjective animal could shuffle the elemental schemata making up its habitual repertoire and try out new combinations, testing them by the consequences associated with each in past experience, it could discover new complex behavioral schemata that are likely to work better.
If the trials were informed by memory of past successes and dangers, the random variation would come with certain emotions attached, including warning of possible dangers in acting on them as well as the attraction of potential benefits.
Dreaming requires long periods when the animal was not otherwise occupied, and that would be provided by regular periods of sleep. Indeed, if dreaming is functional in this way, it may be the function that explains sleep in mammals.
For further evidence of this function of dreaming, see Jonathan Winson's Brain and Psyche: The Biology of the Unconscious.
Acting. The selection of the kind of behavior toward an object is, however, a multi-step process. First, the goal selection system attaches a desire to the object. The animal system of representation must then figure out how to adapt a relevant behavior schema to the world. Finally, something must trigger the command to convert covert motor commands into overt motor commands. That is, it must act on its plan. The systems responsible for the first two steps have been explained, but the decision to act on its plan requires an additional mechanism. When we take up the structure of the mammalian brain, therefore, we will also be looking for a mechanism that makes acting overtly depend on a state of the animal behavior guidance system as a whole.
![]()