![]()
![]() The
origin and detailed structure of the mammalian brain. Assuming that
much of the detailed connections among neurons can be acquired from the structure
of the world by experience, or learning, all that is needed for telesensory
vertebrates to try out the higher level of neurological organization required
for a subjective animal system of representation is a random variation that
sets up the basic circuits depicted in the functional description of the behavior
guidance system. By considering the structures of the non-mammalian and mammalian
vertebrate brains, we will see how it can be accomplished by a relatively
simple, albeit radical, the random variation. That description of the basic
structure of the mammalian brain also reveals the detailed neural mechanisms
that serve the various functions in the subjective animal behavior guidance
system.
The
origin and detailed structure of the mammalian brain. Assuming that
much of the detailed connections among neurons can be acquired from the structure
of the world by experience, or learning, all that is needed for telesensory
vertebrates to try out the higher level of neurological organization required
for a subjective animal system of representation is a random variation that
sets up the basic circuits depicted in the functional description of the behavior
guidance system. By considering the structures of the non-mammalian and mammalian
vertebrate brains, we will see how it can be accomplished by a relatively
simple, albeit radical, the random variation. That description of the basic
structure of the mammalian brain also reveals the detailed neural mechanisms
that serve the various functions in the subjective animal behavior guidance
system.
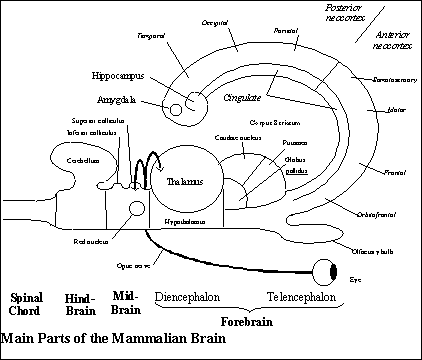
In the non-mammalian vertebrate brain, as we have seen, the three subsystems serving the three subfunctions of the animal behavior guidance system are distributed to three anatomically distinct regions of the brain, the segment of nervous system rostral to the spinal chord, which connects the brain to the body. Those regions are the forebrain, midbrain and hindbrain.
Any behavior guidance system, as we have seen, must register sensory input from the world, select from among incompatible kinds of behavior, and generate the kind of behavior chosen. In the vertebrate brain, sensory input is registered in the midbrain, selection among incompatible kind of behavior is mainly the responsibility of the forebrain, and behavior is generated by the hindbrain. Connections between the midbrain and hindbrain enable them to serve as the telesensory animal system of representation.
The transformation of the non-mammalian into the mammalian brain is often described as the centralization of functions in the nervous system. That is, the three subfunctions of the animal behavior guidance system, which are parceled out to the three anatomically distinct sections of the non-mammalian brain, are all served by one of those sections in mammalian brains, namely, by the forebrain.
It is obvious that the mammalian forebrain serves the output subfunction, since motor output is generated directly from the neocortex, a distinctively mammalian structure in the forebrain. (Motor output from the hindbrain, which generates motor output in non-mammalians, is fully under its control.) Muscles throughout the body are controlled by a somatotopically organized 2 .D array of neurons in the motor neocortex.
Although visual input is already received by the forebrain in non-mammalians (along with the optic tectum in the midbrain), it is joined in mammals by well defined relays to the neocortex for auditory and somatosensory input (all three via nuclei in the thalamus).
Since the subfunction of selecting goals for behavior is already contained in the forebrain, it is apparent that the mammalian forebrain is using its sensory input to generate motor output, thereby serving all three subfunctions. Hence, it is the mammal's entire animal behavior guidance system.
The shift of all three subfunctions from the three distinct parts of the non-mammalian brain to the forebrain resembles the evolution of the vertebrate brain from brainless chordates: just as the rostral end of the chordate neural tube became a brain that took over the control of behavior from reflexes located throughout the neural tube, so the mammalian forebrain takes over the control of behavior from the midbrain and hindbrain. The midbrain and hindbrain structures are still there, but they are slaves to the forebrain (and are omitted from the mammalian diagrams unless they play an essential role).
Furthermore, the neural connections among the subsystems in the mammalian brain are just what would be expected of an animal system of representation with spatial imagination. We have seen functionally how the subjective animal system of representation requires four subsystems (the local image, body image, behavior generator, and memory), and the simplest and most convincing way to show how their functions are served by structures in the mammalian forebrain will be to describe how those mammalian structures evolve from simpler mechanisms in the non-mammalian forebrain. The relevant structures of the vertebrate brain have already been described (see Stage 5. Telesensory animals: Structure of the non-mammalian vertebrate brain).
There are, as already mentioned, two kinds of changes in the non-mammalian forebrain that result in mechanisms for the subjective animal system of representation — the evolution of the neocortex, and a restructuring of the thalamus that redirects the non-mammalian forebrain output back to the (telencephalon of the) forebrain (by way of the thalamus) to form the complete circuits of its higher level of neurological organization. The first change occurs in the telencephalon of the forebrain and the second in the diencephalon.
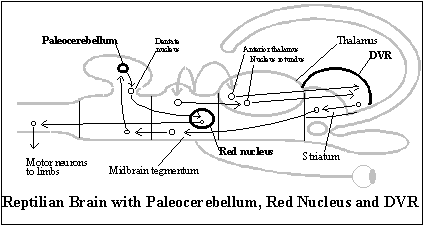
The neocortex evolves from the integration of the DVR and dorsal cortex, the two main telencephalic targets of thalamic nuclei in reptiles. During embryological development in mammals, cells in the telencephalon that would become the DVR in reptiles migrate to the dorsal cortex (the extended flap of neural tube at the diencephalon) and lay themselves out in the dorsal cortex against the outer surface of the mammalian telencephalon. The result is the six layers of cells distinctive of mammalian neocortex. (Compare the diagram of the reptilian brain with that of the mammalian brain.[1]) The evolution of the neocortex adds to the telencephalon a somatotopic representation of the body (the body image) and all the telesensory images that make up the local image.
The thalamus is restructured so that it redirects the non-mammalian forebrain's three kinds of output back to the telencephalon, and this results in the creation of three circuits in the mammalian forebrain. Each circuit is a pathway of neural projections that includes both the thalamus and the neocortex along with special structures that make them different from one another. And each circuit serves one of the basic subfunctions of the animal behavior guidance system. The sensory input circuit serves as local image with the memory in the functional diagram. The behavioral output circuit combines the systems labeled body image and behavior generator in the functional diagram. The third circuit is the affective system, for selecting goals of mammalian behavior, which attaches desires to objects in the local image by activating dispositions in the behavior generator.
Since the neocortex and thalamus are what are linked by the three circuits, these two main changes are connected. Each of the areas of neocortex and nucleus of the thalamus is on roughly the same level of neurological organization as each of the basic systems making up the telesensory animal behavior guidance system, and thus, when they are linked in a three different circuits, each serving one of those subfunctions, there is obviously a higher level of neurological organization. But the best way to see how this basic structure is tried out is to start with the change in the thalamus that gave the neocortex a somatotopic representation of the body to serve as the behavioral output system of the subjective behavioral output system.
The body image. Motor output is generated from the body image, a region of the neocortex that is somatotopically organized. It receives somatosensory input from all parts of the body and it generates the motor output controlling all parts of the body. Like the local image in the posterior cerebrum, the body image is a region of neocortex located in the anterior cerebrum. But somatosensory and motor neocortex represent the body like a picture (or, rather, several pictures), and a somatotopic organization is such a highly improbable mechanical order that the behavioral output system is a good example of a complex structure that seems unlikely to be tried out as a random variation by the mechanism of embryological development. But it most come from somewhere.
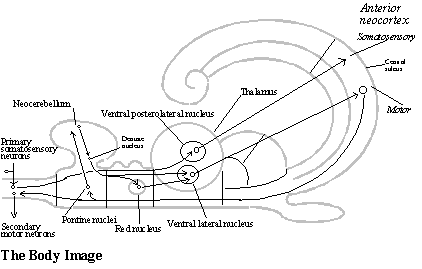
The image of the body in the neocortex cannot come from either of the geometrical structures from which the neocortex evolved, because neither the DVR nor the dorsal cortex have a somatotopic organization. But telesensory animals needed such a somatotopic representation of the body in order to even out motor commands to muscle groups in each region of the body so that whole body behavior would be coordinated. That requires the command neurons to be related spatially to one another corresponding to spatial relations among the parts of the body they control, and that was the function of the cerebellum.
Even before reptiles, there was a rough somatotopic organization in the cerebellum (the archicerebellum), but the evolution of legs required a more distinct somatotopic organization. As mentioned above, it is found in a new region of the cerebellum (the paleocerebellum) and in the red nucleus to which it projects in the midbrain in order to affect motor commands to the limbs.
There is a further development of a similar kind in the mammalian cerebellum. Yet another region is added to the cerebellum (the neocerebellum). The mammalian cerebellum projects to the red nucleus and (joined by a projection from the red nucleus) to the motor neocortex (relayed in both cases by a new nucleus of the thalamus, the ventral lateral nucleus). In other words, the red nucleus now sends motor commands to the limbs by way of a somatotopic representation in the neocortex. (See diagram of the behavioral output system.) This relay of its output to the motor neocortex required a new nucleus in the thalamus, and that may have been the original random variation that is responsible for both the reorganization of the thalamus and the neocortex.
Not only does it locate a somatotopic behavioral output system in the neocortex, but it is also the source of the somatotopic organization of somatosensory input to the neocortex (which is also relayed to the neocortex by a new thalamic nucleus, the ventral posterolateral nucleus).[2] In primitive mammals, such as the hedgehog, somatosensory input and motor output share the same body image in the neocortex, and only later do their images come to occupy adjacent areas of neocortex across the central sulcus.
The local image. The telesensory images making up the local image are on the same level of part-whole complexity as the body image. Both are regions of neocortex receiving highly organized 2-D projections from specific nuclei of the thalamus. As in earlier vertebrates, telesensory input is relayed to the telencephalon by nuclei in the thalamus. Visual input is relayed by the lateral geniculate body to the occipital neocortex, and auditory input is relayed by the medial geniculate body to a region of the temporal neocortex. But the evolution of the neocortex provides a more powerful mechanism for its analysis than in telesensory animals. (See the diagram of the local image.)
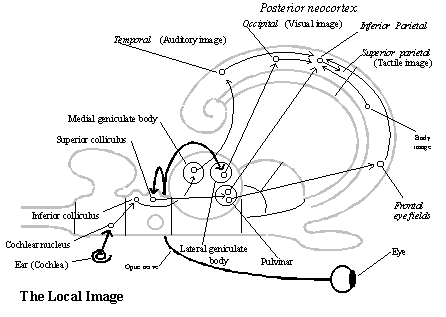
Regions of neocortex all have a similar structure. Thalamic neurons send axons in ordered, 2 .D arrays to certain 2-D areas of neocortex where they synapse with certain kinds of neurons (that is, with neurons in one or another of its six layers). And neurons in target areas of the neocortex send 2 .D arrays of axons to three kinds of targets in the forebrain: directly back to thalamic nuclei, to other neocortical areas, and to the corpus striatum and its special projection to certain thalamic nuclei (behavior generator). These connections are determined genetically, by instructions from the multicellular biological behavior guidance system. It is a good example of the more complex structures that can develop from the neural tube in deuterostomes: neurons in the thalamic nuclei and their neocortical targets are born with molecular labels by which they can somehow recognize their own locations in their own 2 .D array and seek out corresponding locations in other 2 .D arrays.[3]
In the case of telesensory images, all the 2 .D arrays in the thalamus and neocortex are usually processing sensory input from the same objects in the local scene at any given moment, but following genetic instructions to analyze it in different ways. With evolution by reinforcement selection, therefore, they can use the results of one another's analysis to confirm their own conclusions, thereby improving the efficiency of them all.[4] Jointly, they are a subsystem with a 2 .D representation of the object at a distance.
The local image is, however, a more encompassing 2-D representation of the current scene, which includes all the currently perceivable objects relative to the body. The local image is constructed by an interaction between the body image and specific telesensory representations. The local image is constructed in a region of neocortex (the inferior parietal lobe) adjacent to the visual and auditory analysis areas, on one side, and the body image, on the other, and it receives association fibers from both sides. (See diagram of the local image.) The image of the body in the motor and somatosensory neocortex projects association fibers providing information about the current bodily condition (its stance and motion), and the parietal area uses it to locate objects of telesensory images in the scene relative to the body by taking account of the direction in which the head and ears are pointed when they are received.
The eyes are, however, a special case. Their fixations are controlled by the frontal eye fields (an area of frontal neocortex, anterior to the motor cortex, which we are counting as part of the behavioral output system). The frontal eye fields also report their oculomotor commands to the inferior parietal lobe (via association fibers), where an interaction with the visual neocortex (in the occipital area) constructs a larger representation of the scene (the local image) from the visual images that show up in each fixation.[5] (See diagram of the local image.) The already constructed local image guides the fixations of the eyes to objects in the scene; that is apparently the function of a highly unusual, ordered 2-D projection of fibers to the frontal eye fields from the thalamic nucleus (pulvinar) that serves the inferior parietal area.[6] The local image is not, therefore, just a collection of telesensory images, but an ordering of them that indicates the direction and distance of each from the body, or what serves as representation of the current local scene.
Touch is a new sensory modality, which depends on a certain kind of somatosensory input to the body image. As we have noticed, somatosensory input is relayed (by the posterolateral nucleus of the thalamus) to the somatosensory neocortex (across the central sulcus from the motor neocortex), where reports from joint and muscle receptors supplies feedback about the effects of motor commands from the body image. But it is also, in effect, a new sensory modality, because input from skin receptors (of various kinds) is combined with the representations of the scene and the body (in the superior parietal area served by the lateral posterior thalamic nucleus) to yield information about other objects in the current scene (though this does not yield much information about them except location until the evolution of manipulative animals). (See diagram of local image.) Since the objects are distinct from the body, but located not at a distance for it, touch might be called an "extrosensory modality."[7]
The evolution of the neocortex, in sum, accounts for one part of each of the two basic subsystems of the subjective animal system of representation, the sensory input and behavioral output systems. Almost every area of neocortex receives a projection from one or more thalamic nuclei (and sends fibers back), but the body image is located in the anterior neocortex, and the 2 .D telesensory images of the objects in the scene and their combination as a local image is located in the posterior neocortex. But these regions of neocortex act on the thalamus in two different ways, each of which makes the neocortex a link in a complete circuit. The rest of each circuit is the other part of the sensory input and behavior output systems represented in the functional diagram of the subjective animal system of representation, and that is the part of each circuit that is provided by the other major change in the mammalian forebrain, the redirection of the output of the non-mammalian forebrain back to the telencephalon by relays in the thalamus. (The same also holds for the goal selection system, the third part of an animal behavior guidance system, as we shall see.)
We have seen that there are three pathways of output from the non-mammalian forebrain: the ancient olfactory pathway, the striatal pathway, and the hippocampal pathway. And each is responsible for a new, complete circuit in the mammalian forebrain. Indeed, the functions are parceled out to these circuits according to the three subfunctions of any behavior guidance system.
The striatal pathway becomes part of the behavior generator which works together with the body image to serve as the behavioral output system.
The hippocampal pathway becomes the memory which works together with the local image to serve the sensory input subfunction of mapping the territory.
And the olfactory pathway becomes the goal selection system which works together with both frontal and posterior areas of neocortex to serve the subfunction of selecting goals,. We will put the olfactory pathway aside for now and take it up only after detailing the subjective animal system of representation, because the subjective animal behavior guidance system is what results when olfactory pathway is added to the subjective animal system of representation. Let us begin with the hippocampal pathway and how memories are recorded.
The memory. The mammalian memory is a forebrain circuit that results from redirecting the hippocampal output of the non-mammalian forebrain back to the telencephalon by way of another new thalamic relay (the anterior nucleus) in the diencephalon. Its target is, however, a new region of neocortex, the cingulate area, which runs parallel to all the other areas of the neocortex at the medial extreme of the neocortex (including the local image, body image, and other regions of frontal neocortex where behavioral schemata are located. See diagram of the memory.) This provides a new, complete circuit, because association fibers connect the cingulate area with all the areas of neocortex running parallel to it, and those areas all have association fibers to (a staging area in the temporal lobe that projects to) the hippocampus. Since the hippocampus receives information about the "images" in 2-D arrays throughout the neocortex, it is what enables the mammalian brain to learn from experience with the three dimensional structure of the world how to use the input about the body condition (association fibers from the behavioral output system to the parietal cortex) to construct the representation of the local scene and how to label such representations by their locations relative to one another in three dimensional space as it forms new memory.
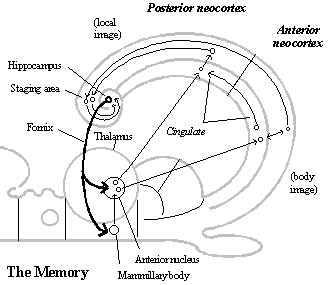
The basic mechanism involved in the formation of memories is the strengthening of synapses of association fibers connecting sets of neurons in various regions of the neocortex that are firing at the time, so that later, when enough of them fire together again, it makes the others in the group fire as well. Although synapses are strengthened anyway among 2-D arrays in nearby areas of neocortex that are closely interconnected,[8] the hippocampus-cingulate circuit connects, as memory groups, neurons from all areas of neocortex that are active at the moment.[9] This connects images of the same object from different sensory modalities as the same object and images of various objects in the current scene as a local image so that they can be called up together as units. But it is necessary to have input about the current bodily condition to label telesensory images by the behavior involved in perceiving them in the scene. That is provided by association fibers from the behavioral output system to the inferior parietal neocortex, and if input from the bodily condition is not what is combined in the memory groups for the local image, it must be the relevant groups of neurons of the behavioral output system itself.
Given how the memory system (the hippocampus-cingulate circuit) works, it is required to form such cortex-wide memories. But it is not involved in recalling memories. Recalling memories is simply a matter of making the whole group fire together by enough of its members firing for some reason.[10]
Indeed, at an earlier stage of embryological development, as mentioned above, the memory circuit is responsible for acquiring the behavioral schemata by which overt inquisitive behavior can construct local images and similar covert behavior can extract information about the locations of objects in the local science from local images called up from memory. The memory circuit provides the “natural selection” for a contained form of reproductive causation in which the “reproducing organisms” are connections among neurons serving as behavioral schemata, neurons representing the bodily condition, and neurons serving as telesensory images, and these connections evolve as parts of schemata that the behavior generator can use, as inquisitive behavior, to construct local image, or as covert behavior, extract information about them. The correlations between telesensory input and bodily condition that hold for objects of every kind in any local scene depend on the structure of space (and the structure of the body driven by the brain), and the memory circuit enables the brain to acquire the general connections between cells in the local image and body image so that behavior schemata for constructing and using local images can evolve by the natural selection of a reinforcement that strengthens the connections when they are successful, for example, when the animal can find objects it desires in the scene. Thus, the structural cause of the capacity to record and use local images is internalized from the structure of the spatiomaterial world by a form of reproductive causation that occurs as a later phase of the process of the embryological development of the brain.
There is evidence, by the way, that the schemata for this kind of behavior is derived from experience. When human subjects are fitted with glasses that reverse right and left or up and down in the visual field, there is, after a long period of awkward attempts to compensate, a dramatic change in which subjects report that everything "suddenly" seems oriented in the "right" direction again. As the memory circuit lays in new connections for behavioral schemata to construct and use local images with the reversing glasses and they take over, one's sense of direction changes.
The memory circuit also has a built-in structure that makes it possible to connect local images in sequences corresponding to the direction of locomotion and turning relative to them, though linking them in such sequences depends on the cingulate neocortex functioning in an additional way. When the cingulate neurons in a memory group fire, so do the associated images in the adjoining regions of neocortex, and vice versa. The means that the cingulate neocortex is in a position to determine the local images called up in sensory input system. Thus, in order to serve as the memory for spatial imagination, all the cingulate gyrus needs is the capacity to link local images together in long sequences and to be able to switch from one such sequence to another when their local images overlap.
The posterior cingulate gyrus is in a position to serve as the memory for spatial imagination, because the posterior cingulate receives input about the bodily condition, overt and covert, from the anterior cingulate gyrus. The anterior cingulate gyrus is connected to the body image and the behavioral schemata in the behavior generator in a way that is similar to the connection between the posterior cingulate gyrus and the local image, except that the influence runs the opposite direction. The body image and behavioral schemata project to the anterior cingulate gyrus, and the anterior cingulate gyrus is an (agranular) motor neocortex like the body image and behavioral schemata, except that it has not effect on overt motor output and projects to the posterior cingulate gyrus.
Since the body image contains information about how the body moves and turns overtly in the scene, the connections it makes with the posterior cingulate gyrus would become labels that are recorded with the sequences of local images. And since the behavioral schemata contain the same kind of information about covert behavior, and it is projected to the posterior cingulate gyrus in the same way, those connections can be used as labels to call up the local images according to how it move and turns covertly in relation to the images called up in memory.
As sequences of local images become richly interconnected, the mammal acquires a map of its territory by which it can travel almost any route. It is the structure of space that ensures that such interconnections of sequences can accumulate without contradictions. This internalization of the structure of space is an example of the a contained form of reproductive causation that takes place in neurological development as a result of reinforcement selection. The memory labels required for constructing and exploring memory maps are part of the behavior schemata that evolve gradually toward greater efficiency in satisfying desires, in the first instance, getting the animal where it wants to go.[11]
The behavior generator. In reptiles, the DVR is the main source of input to the striatum (which is one pathway of non-mammalian forebrain output to the midbrain), and thus, since those DVR cells migrate in mammals to the dorsal cortex and combine as the neocortex, the neocortex becomes the main source of input to the striatum (called the corpus striatum or basal ganglia). In fact, all areas of the neocortex project to the corpus striatum, including the local image, the body image, behavioral schemata in the frontal neocortex, and the cingulate neocortex. But the output of the corpus striatum, which in reptiles was projected to the midbrain as motor commands, is mostly redirected in mammals back to the neocortex by way of a relay in the thalamus (the ventral lateral and ventral anterior nuclei). The targets of the corpus striatum are all in the frontal neocortex (in the anterior forebrain), including both the body image and the behavioral schemata. This new circuit is the behavior generator. But there are two channels through this circuit, which were not distinguished in the functional diagram except for the ability to generate behavior either overtly or covertly.
Part of the corpus striatum's output is relayed by the (ventral anterior) thalamus to regions throughout the frontal neocortex where the behavioral schemata are located. This part also projects to a region called the “premotor cortex” (an anterior part of Brodmann area 6), which is the behavioral output system for covert behavior. It also projects to the frontal eye fields, which controls the fixation of the eyes on objects in the local image.
The other part of the striatum’s output is relayed by the (ventral lateral) thalamus more specifically to the motor neocortex itself, the part of the body image that is responsible for overt behavior. This includes both the motor neocortex (Brodmann area 4) the supplementary motor neocortex (an area of neocortex adjacent to the motor neocortex which modifies motor commands directly, which is part of Brodmann area 6.)
The motor neocortex (the body image from which overt behavior in generated) also receives a projection from the ventral lateral thalamus that relays the output of the cerebellum (where motor commands are evened out spatially and temporally in a detailed way). The motor commands actually sent to the body depend, therefore, on the cerebellum as much as the circuit through the corpus striatum and ventral lateral thalamus. But the cerebellum is controlled by these circuits through the striatum and thalamus, because the cerebellum follows motor commands from the body image, including both parts, Brodmann areas 4 and 6. (The premotor commands, a part of the body image that generates covert behavior, enables the cerebellum to anticipate the next overt motor commands.)
The corpus striatum has just the connections required of a subsystem whose function is to generate behavior in relation to the object at a distance. It receives 2 .D arrays of fibers from all areas of the neocortex, including the local image in the posterior neocortex, on the one hand, and the behavioral schemata and body image (both premotor and motor areas) in the anterior neocortex. But it projects only to the behavioral schemata and the body image in the anterior neocortex. (See diagram of the behavior generator: overt behavior.)

The local image sends fibers almost exclusively to a distinct part of the corpus striatum called the caudate nucleus. The caudate nucleus apparently compiles an image of the object situated in the current scene from all parts of the posterior neocortex, because 2 .D arrays of posterior neocortex that are connected by association fibers tend to converge on the same areas of the caudate nucleus.[12]
Similarly, the body image send fibers almost exclusively to a distinct part of the corpus striatum, called the putamen. The putamen compiles an image of the body from all areas of the anterior neocortex, including the frontal areas containing behavioral schemata and the body image, for it merges 2-D arrays from parts of neocortex that are closely connected by association fibers. With behavioral schemata contained in the frontal neocortex, the putamen has instructions by which to generate a specific kind of behavior (depending on which desire predominates at the moment).
A third distinct part of the corpus striatum, called the globus pallidus, combines the compiled image of the body with the compiled image of the object according to the selected schema to produce detailed motor commands for each part of the body as needed to adjust its behavior to the object in space.[13] The globus pallidus can generate behavior, because it projects by way of nuclei in the thalamus to areas of premotor and motor neocortex that are responsible for motor commands to all parts of the body.
For the animal system of representation to have a spatial imagination, however, the behavior generator must be able to generate motor commands in relation to objects in the scene without the body actually acting on them. And the corpus striatum does, indeed, have connections that make it possible to generate behavior covertly rather than overtly. There are two distinct channels by which the corpus striatum projects to the thalamus (and from there back to the neocortex, which completes a circuit because of the cortex’s projection to the corpus striatum). (Both originate in the internal segment of the globus pallidus, the third part of the corpus striatum, where both the local image and body image are represented).
One channel (by way of the ansa lenticularis) projects to the ventral anterior nucleus of the thalamus which relays it to the premotor and other frontal areas of neocortex where behavioral schemata are contained. These areas of neocortex project in turn to the anterior cingulate gyrus, whose connections with the posterior cingulate gyrus enable covert behavior to affect the sequences of images in memory.
The other channel (by way of the lenticular fasciculus) is relayed by the ventral lateral thalamic nucleus to the supplementary motor neocortex, which lies alongside the motor neocortex and generates overt behavior. (The ventral lateral nucleus also relays the detailed motor instructions from the cerebellum and red nucleus, but that is a later effect of this circuit for overt behavior through the striatum and thalamus, because its motor commands are what the cerebellum uses to work out the detailed motor commands.) These areas of the body image also project to the anterior cingulate gyrus, where they can establish labels for the sequences of local images in spatial imagination.
Both channels of the are active when behavior is overt. The covert route is never suppressed, because it is the circuit that contains the behavioral schemata and uses them to set up complex, multi-step motor commands in advance of acting. But when the channel to the motor neocortex is suppressed, behavior is generated covertly, as imagination, because the anterior cingulate gyrus still receives the projection from the behavioral schemata and premotor cortex. That is enough information about locomotion and turning to call up memories of local images by the labels that were established when both overt and covert parts of the body image were being projected to it.
The subjective animal’s behavioral output system has, therefore, two body images. Both are used for generating overt behavior, but one can be used for generating covert behavior. Covert behavior is the motor output in the cingulate gyrus, which acts, not on the actual world by way of the body, but on the world as represented in memory by way of covert behavior. Since the anterior cingulate gyrus projects to the posterior cingulate gyrus in much the same way that the motor neocortex (and associated areas) project to the parietal and sensory neocortex containing the local image, it is not surprising that the memory circuit can register images according to the behavior that causes them, that is, by labels. Nor is it surprising that covert behavior can use those labels to call up the images from memory as imagination.
The difference is that telesensory images are registered according to input from the overt bodily condition so that they become a local image, which can be recorded as a unit in memory, whereas local images are registered according to input from the overt locomotion so that they become linked in a sequence as a map of the territory. One occurs in the non-cingulate region of the neocortex, the other occurs requires the cingulate area. Likewise, when a local image is remembered, its individual telesensory images can be called up by input from covert behavior in the non-cingulate neocortex, whereas other local images connected to it can be called up by input from covert locomotion in the cingulate gyrus.
This parallel suggests that labels can be established in both cases by the memory circuit working as a form of reinforcement selection for the evolution of behavior schemata. The labels are simply connections of the behavioral schemata with the rest of the brain that are reinforced as parts of those schemata when behavior is overt, and thus, since the behavioral schemata can be used to generate the same behavior covertly, they automatically have labels by which to call up the relevant images from memory.
What is crucial to internalizing the structure of space as spatial imagination about the consequences of locomotion and turning is the capacity of the cingulate gyrus to link complex memory groups in sequences. That is what the non-cingulate neocortex lacks. Sequences of images are not needed to construct local images, because they can be accessed in any order by the appropriate bodily condition, turning the head, eyes, etc. But it is the use of sequences, and their being projected to the behavior generator (via the caudate nucleus of the corpus striatum) as time passes, that makes it possible to imagine locomotion and motion. And that is the key to their being an understanding of the structure of space. This is a capacity to see the actual locations of objects against the background of what is possible by motion, which is to understand the structure of space.
The subjective animal’s map of the world becomes an understanding of space simply because the same kinds of locomotion and turning have the same kinds of effects relative to any set of local images. That is because the world from which those sequences were recorded is a world of objects that are actually related by being contained by space. Thus, experience with the world gives the subjective animal is understanding of the general effects of the motion and direction of motion of an object on its spatial relations to other objects. It is, of course, an intuitive understanding, not a linguistic understanding. But it is nonetheless an understanding by which it can foresee what will happen and understand what has happened.
More generally, however, the power of the memory circuit as a form of natural selection on behavioral schemata working by the reinforcement of synapses to form memory groups is broader. Assuming that during the process of development, the whole range of behavioral schemata that are possible because of the genetically provided basic schema are tried out, gradual evolution will occur, and the schemata will becomes increasingly powerful. Each behavioral schema will come to be as effective as possible in controlling some relevant condition, that is, generating behavior is some kind of situation to satisfy desire. That is actually the result of the gradual evolution of that schema, since it is made up of different motor commands as parts, and it is by alterations in them that variations are tried out. But different behavioral schemata will also be fit together as harmoniously as possible as parts of behavioral routines that satisfy desire. And new schemata will be added as long as they control additional conditions that affect their reproduction. Thus, learning from experience that provides subjective animals with behavioral schemata for many kind of behavior. The schemata that serve as structural causes for spatial imagination is just the most important kind of behavior, given the nature of the world in which animal find themselves and their need to act on objects in space.
This theory of imagination as covert behavior offers a plausible explanation of schizophrenia. Hallucinations that are not recognized as hallucinations (usually auditory hallucinations) are the main symptom of schizophrenia. They make it difficult for schizophrenics to tell the difference between fantasy and reality. It is known that schizophrenics often show a lack of activity in the frontal neocortex when hallucinating (apparently due to an unusual level of the neurotransmitter, GABA, in the region). Those areas of neocortex contain the behavioral schemata that control the anterior cingulate gyrus, whose effect on the posterior cingulate is to call up images from memory, and thus, it is possible that schizophrenia is a condition in which covert behavior is generated without control by behavioral schemata in the frontal neocortex. Since the images are not called up by activating some behavioral schema, as in normal behavior and imagination, they are not part the process of carrying out an intention, and thus, they seem to originate from the world, that is, to be real.
The goal selection system. Not only does reproductive causation in neurological development require reinforcement, but once animals have acquired a map of the territory and an understanding of the structure of space, they still need a mechanism to choose which goal to pursue in relation to which objects. These functions are served by the redirection of the third output of the non-mammalian forebrain back to the telencephalon. The redirection of the olfactory pathway back to the telencephalon makes it the goal selection system of the mammalian behavior guidance system.
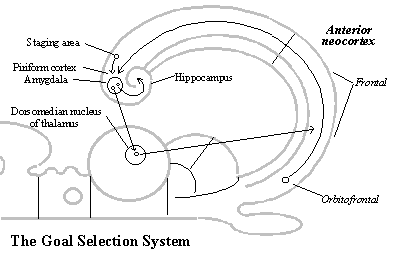
The olfactory pathways involve, as described in the non-mammalian vertebrate brain,[14] the amygdala and its many connections with the hypothalamus (and septum). This is still the seat of desire in mammals. The amygdala is mainly responsible for arousal, fear and anger, whereas the hypothalamus is responsible for hunger and sexual desire.
In mammals, the output of the amygdala (and indirectly the hypothalamus) is redirected by a thalamic nucleus (the dorsomedian nucleus) back to the behavioral schemata in the frontal neocortex (especially, the orbitofrontal area). This system selects which of the schemata for kinds of behavior that are built into the frontal neocortex (genetically or by learning) will guide the corpus striatum in generating behavior relative to the object in the local image.
The goal selection system is a complete circuit, for there are projections from the orbitofrontal area back to the amygdala (via the ancient piriform cortex surrounding the amygdala) which make sure that there is no confusion about which object is the object of desire.
Furthermore, the goal selection circuit is in a position to record and read desire labels with memory images. The amygdala has a projection to the hippocampus by which it can attach desire-labels to objects (for example, when the hypothalamus reports to the amygdala that an object satisfies hunger or sexual needs), and it can read the desire-labels already attached to objects, because it receives a projection from the staging area by which all areas of neocortex reach the hippocampus. (See diagram of the goal selection system.[15])
A desire about an object sets a problem for the subjective animal system of representation to solve. Covert locomotion can, for example, explore its map and figure out a new route to an unperceived goal when its path is blocked or use its representation of the current scene to figure out how to approach or avoid some object. But a spatial imagination is only part of what is possible for a system that uses a somatotopic representation of the body to generate behavior by its parts, like the body image. Instead of having to evolve new instincts, it can learn new kinds of behavior involving different combinations of motor commands to parts of the body. And mammals can, in principle, figure out how to use such complex routines in the current scene by imagining variations on them and using memory images that are called up to anticipate whether the consequences will satisfy its desire. At least, this is the direction of subsequent evolution, as we shall see.
Acting. Once the problem is solved and a plan of behavior has been selected, the imagined behavior must be generated overtly. Covert behavior depends on one of the pathways through the behavior generator (the one from the corpus striatum via the ansa lenticularis to the ventral anterior nucleus of the thalamus and back to the frontal neocortex and premotor area). The behavioral schema that has been set up in imagination can be generated overtly by sending the same motor commands in relation to the perceived object along the other pathway as well (via the lenticular fasciculus to the ventral lateral nucleus back to the motor cortex).
What controls whether covert behavior is generated overtly or not is the centromedian nucleus of the thalamus. It projects to the putamen, the part of the corpus striatum that receives the schema from the frontal neocortex for the kind of behavior to be generated. The centromedian nucleus is located within the medullary lamina of the thalamus (separating its medial and lateral subdivisions), and since the thalamus is what synchronizes activity throughout the neocortex, that is a position from which it can to tell when an equilibrium is reached in systems throughout the brain. The intralaminar nuclei of the thalamus receive fibers from all parts of the neocortex, the brainstem, cerebellum and spinal chord.[16] (See diagram of the mechanism of acting.)
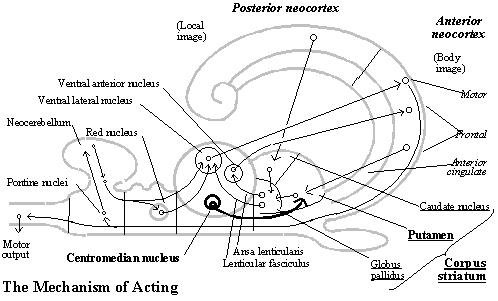
In the transformation of the non-mammalian into the mammalian brain, therefore, we find all three subsystems required for the subjective animal behavior guidance system (sensory input, behavioral output, and goal selection systems). Each involves a distinct circuit through the thalamus and neocortex, which allows neurons in multiple 2-D arrays throughout the circuit to be coordinated and aligned with one another. That puts each subsystem on a higher level of neurological organization than the non-mammalian vertebrate brain. Moreover, in the sensory input and behavioral output circuits we find both subsystems required according to our functional diagram for the subjective animal system of representation with spatial imagination (that is, local images and memory in the sensory input system and behavior generator and body image in the behavioral output system).
As we have seen, such a complex system is possible, at least in chordates (with the nervous system set up in a separate neural tube), because the higher level of neurological organization can be generated by a couple of relatively simple, albeit radical, modifications in the non-mammalian vertebrate brain, and neurological development involves a contained form of reproductive causation that internalizes the spatial structure of the world as the detailed structural causes of spatial imagination. Since the subjective animal behavior guidance system is also functional, we infer by the nature of reproductive causation that this stage of animal evolution in inevitable.
In the next section, we will consider the empirical evidence of the subjective animal stage of evolution. Though empirical confirmation of necessary truths is not necessary, it is relevant, because empirical evidence can falsify the foundation of ontological philosophy.
That will leave one final conclusion to be drawn, about the unity of consciousness in mammals.
It is appropriate to think of the mammal as a subject, since its spatial imagination gives it an understanding the structure of space and the regularities about motion and interaction for which it is responsible as an ontological cause. By seeing the actual locations of objects against the background of what is possible by motion, the subject sees the world as a world of objects in space. Even its own body is seen as located in space alongside other objects. But the mammal is not just a subject. It is a conscious subject.
When combined with the explanation of the basic nature of consciousness (that is, phenomenal properties as intrinsic essential natures of bits of matter), the structure of the mammalian brain also explains the unity of subjective consciousness, that is, why all the sensory qualia appear to a single subject at one time. There is a phenomenal appearance to the world of object in space and the body by which the subject acts in that world. Nor do the objects that subjective animals experience as located in space appear merely as object in space, because they also feel desires of various kinds about them. We will return to the unity of consciousness after considering the empirical evidence about the gradual evolution of mammals.
![]()