![]()
![]()
 5. Telesensory animals. Telesensory animals are multicellular animals with a centralized nervous
system in which the three basic subfunctions of the animal behavior guidance
system are served by entire systems of neurons. They include most invertebrates
along with non-mammalian vertebrates, and after explaining why the basic structure
of the invertebrate nervous system limits their subsequent evolution, this
chapter describes the structure of the vertebrate brain in detail and traces
the actual evolution of vertebrates on earth. But in order to deduce this
stage of evolution, it must first be shown to be inevitable.
5. Telesensory animals. Telesensory animals are multicellular animals with a centralized nervous
system in which the three basic subfunctions of the animal behavior guidance
system are served by entire systems of neurons. They include most invertebrates
along with non-mammalian vertebrates, and after explaining why the basic structure
of the invertebrate nervous system limits their subsequent evolution, this
chapter describes the structure of the vertebrate brain in detail and traces
the actual evolution of vertebrates on earth. But in order to deduce this
stage of evolution, it must first be shown to be inevitable.
![]() Inevitability
of telesensory animals. According to this ontological explanation
of evolution as a global regularity, the evolution of multicellular animals
with nervous systems at the telesensory level of part-whole complexity is
inevitable, if it is both possible and functional.
Inevitability
of telesensory animals. According to this ontological explanation
of evolution as a global regularity, the evolution of multicellular animals
with nervous systems at the telesensory level of part-whole complexity is
inevitable, if it is both possible and functional.
![]() Possibility.
To be possible, the telesensory level of neurological organization must fall
within the range of random variations being tried out in the evolution of
somatosensory animals. And it is clearly possible, because somatosensory animals
already have already evolved the mechanism of embryological development to
serve as their biological behavior guidance system, and they have already
evolved neurons for transmitting and regulating the signals from sensory input
that caused behavioral output. Thus, all that was required for telesensory
animals to evolve was a higher level of part-whole complexity in the organization
of the nervous system, that is, a centralized nervous system in which neurons
are organized into systems that can serve the basic subfunctions of an animal
behavior guidance system with an animal system of representation.
Possibility.
To be possible, the telesensory level of neurological organization must fall
within the range of random variations being tried out in the evolution of
somatosensory animals. And it is clearly possible, because somatosensory animals
already have already evolved the mechanism of embryological development to
serve as their biological behavior guidance system, and they have already
evolved neurons for transmitting and regulating the signals from sensory input
that caused behavioral output. Thus, all that was required for telesensory
animals to evolve was a higher level of part-whole complexity in the organization
of the nervous system, that is, a centralized nervous system in which neurons
are organized into systems that can serve the basic subfunctions of an animal
behavior guidance system with an animal system of representation.
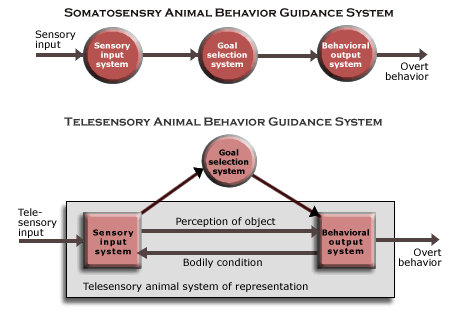
Neurons generating motor output must be parts of a single system, in order for animal behavior to include locomotion in relation to objects at a distance. But since local reflexes in somatosensory animals could already recruit one another to generate whole body behavior, control of locomotor behavior could be supplied by neurons that controlled all the local reflexes.
However, in order to guide locomotion relative to distant objects, sensory organism must be able to detect objects located at a distance from the body, and since that means using proximal input to extract information about distant objects, neurons must be organized on a higher level of part-whole complexity in space. There are only certain kinds of effects on the animal’s body that can be used: electromagnetic radiation, vibrations in the medium or substratum, and kinds of molecules encountered. But since somatosensory animals have already evolved neurons that respond to such effects, it is just a matter of organizing them into systems of neurons whose joint output depends on the distant objects causing the proximal input.
In the case of vision, for example, photo-sensitive neurons must be arranged in a two dimensional array (2 .D array). Not many such neurons are required for the compound eyes of insects, but in camera-like eyes. extremely large 2 .D arrays of neurons are required to register images focused on them by a lens. Hearing requires only neurons that are sensitive to mechanical disturbances, but to discriminate the direction and tone of the sound, they must compare the disturbances of many neurons at once. (Hearing evolve in vertebrates from the lateral line in fishes, which is basically a series of sensors of mechanical disturbances along the side of the body.) Though somatosensory animals already have cells sensitive to kinds of molecules (taste receptors) by which they choose which objects to ingest, olfaction requires a large variety of neurons testing many different aspects of the shapes of molecules at the same time. In each case, multiple neurons are organized as a whole system, and the effects of its geometrical structure are as important to the function of the telesensory organ as the kind of neuron of which it is composed.
These systems of neurons had to be organized as parts of centralized nervous system in order to use telesensory input to guide locomotion (and to select the appropriate kind of behavior as an animal behavior guidance system). And as we have seen, in order to construct an animal representation for guiding behavior in relations to distant objects, brain mechanisms must connect input from telesensory organs with input from their bodily condition, such as the orientation of the body and head and its locomotion. But since the mechanism of embryological development had evolved to construct structures of eukaryotic cells, there is no reason to doubt that the telesensory nervous system is possible.
![]() Functionality.
To be functional and begin another stage of evolution, however, such a higher
level of neurological organization had to enable telesensory animals to control
an entire range of relevant conditions that were out of reach for animals
at previous stages.
Functionality.
To be functional and begin another stage of evolution, however, such a higher
level of neurological organization had to enable telesensory animals to control
an entire range of relevant conditions that were out of reach for animals
at previous stages.
Though it is clear that the ability to use telesensory input would make them more powerful than somatosensory animals, it may not be clear that this power was out of reach for all previous animals, because animals at lower biological levels move about and have mechanisms to guide their locomotion.
Magnetosomes in bacteria, for example, tell which direction to move in order avoid the toxic effects of oxygen-rich water near the surface. And animal-like protists, such Paramecia, have eye spots by which they can orient their motion relative to light.
However, even if this is the capacity to guide locomotion relative to distant objects, it would make telesensory animals inherently more powerful, because they would be guiding the locomotion of a multicellular animal, which is already inherently more powerful than single-celled animals.
In any case, these structures in animal-like protists are not telesensory organs, much less an animal systems of representation, because they do not detect objects at a distance in space. They merely orient locomotion in some ambient field. In these animals, locomotion is just a response to light built into the organism’s structure by the biological behavior guidance system, like reflexes in somatosensory animals.
Since telesensory animals are both possible and functional, this ontological explanation of revolutionary evolution implies that their evolution is inevitable. Telesensory animals would evolve on any planet in a spatiomaterial world like ours where stages of evolution could unfold at all. But since they begin another stage of evolution, reproductive causation also implies that their basic power to guide locomotion in relation to distant objects would evolve in ways that enable them to control additional conditions affecting reproduction. This prediction is confirmed by the main accomplishment of this animal stage of evolution, namely, the evolution of nervous systems with the capacity to map the relations of salient objects in their territory.
Maps. Telesensory animals would be even more powerful, if besides guiding locomotion in relation to objects currently being detected objects at a distance, their animal representations enabled them to guide locomotion in relation to currently undetected objects. Since telesensory nervous systems must already be able to generate more or less complex instinctive behavioral routines in relation to objects in space in order to generate the various kinds of behavior needed to control relevant conditions, only relatively modest modifications of their neurological mechanisms are needed to represent the locations of unperceived objects.
For example, the capacity to keep track of how far the animal moves in each direction relative to the sun or magnetic north would enable it to return to a nest by "dead reckoning" (so that once it is in the vicinity of its nest it could guide its locomotion relative to objects recognized from current telesensory input). Gallistel (1990) has marshaled abundant evidence of such mechanisms in telesensory animals. Desert ants, for example, find their way back to their hive by keeping track of how far they go in any direction relative to the sun or wind. Only when they get in what should be the vicinity of their hive do they orient their locomotion by telesensory input from objects in space. (Gallistel 1990, pp. 59-65.)
The use of large-scale maps by social insects like honey bees is undeniable. Returning foragers direct other members to energy sources by performing a figure-eight dance in relation to the azimuth of the sun that successfully communicates both the direction and distance to other bees. Gallistel shows how these feats could be explained rather simply, assuming that neural mechanisms can represent distances and directions of locomotion and perform quantitative computations on them.
Such telesensory level animal systems of representation are capable of rather sophisticated navigational feats. When honey bees, for example, are released in a region with a known bearing and distance from their hive, not only can they return to their hive, but they can also fly directly to another, unseen energy source with a different bearing and distance from their hive when they have recently been feeding on it and that is the only way to obtain energy (Gallistel 1990, pp. 131-140).
Such territorial maps suggest how much power can evolve in telesensory animals because their animal behavior guidance system contains a system of representation. But to call a mechanism a system of “representation” is to say that there is a correspondence between its representations and what they represent, and it is relevant to consider how such a correspondence is constituted by the animal system of representation, because it is the beginning of an explanation of the nature of truth that will hold of rational beings like us as well.
In the case of the simplest animal systems of representation, the correspondence is basically geometrical. Spatial aspects of behavior are adapted to spatial aspects of the world so that the behavior imposes a geometrical structure on the thermodynamic flow in the region, and what makes that possible is an implicit correspondence between the representation of the object in the animal system of representation and spatial aspects of the world. The representation of the object in the sensory input system has a geometrical structure that corresponds to the relations of objects in space because of how sensory input is recorded together with input from the current bodily condition.
Telesensory organs, like the eye, provide information about the locations and kinds of objects, not only because of the interaction between the eye’s geometrical structure and other objects in space (by way of photons), but also because that telesensory input is contingent on the direction in which the eye is pointing and the head is oriented. Telesensory input is registered in a way that reflects the bodily condition at the time, and since the animal representation can trigger behavioral dispositions that will approach or avoid objects at a distance, it can be used to guide locomotion in relation to them.
The basically geometrical nature of the correspondence between the animal representation and the locations of objects relative to the body solves a problem that functionalist explanations encounter when they try to explain correspondence with nothing but causal connections between input and output within the organism. The correspondence is not just a constant conjunction between telesensory input and the object in space that is involved in reference, as Fodor seems to mean by calling it a “casual connection,” but an isomorphism between a structure in the brain and the geometrical structures about the locations of objects in the space around the telesensory animal. (See Epistemological philosophy of causation: Rational causation: Psychology: Naturalism.)
In the case of maps of the spatial relations of unperceived objects in the territory, the correspondence depends on a more complex geometrical structure than is required to represent the locations of currently detected objects in space, because it involves two phases.
First, since the animal representation of the object functions as a map, telesensory input must be registered in a way that reflects how far the animal must go in certain directions relative to the sun or magnetic north to get from one recognizable location to another. (This is a form of inquisitive behavior.)
Once the map has been constructed, the animal can, during the second stage, use the isomorphism between those brain states and salient objects in the territory to guide its behavior in relation to undetected objects in space.
The construction and use of maps are two different structural global regularities generated by the structure of the nervous system. During the map-making phase, the nervous system acquires an additional geometrical structure from the world, and during the second phase, it uses that unchanging geometrical structure to guide its behavior. The world itself is, therefore, partly the source of the structural cause by which animal behavior imposes a geometrical structure on the thermodynamics flow of matter in the region toward evenly distributed heat. The use of its map enables the animal to anticipate what will happen as it moves in various directions so that it can choose among them according to the kind of behavior that would control relevant conditions. This geometrical fit of its behavior with the environment would be quite mysterious, except for the structural causes involved, including the first phase of map-making behavior that sets up the correspondence involved in representing the locations of salient objects in its territory.
![]() The
Inferiority of Invertebrates. The evolution of telesensory animals
in the direction of natural perfection for organisms of their kind is necessary
for the evolution of subjective animals, but it is not sufficient. The telesensory
stage must make it possible for the mechanism of embryological development
(the biological behavior guidance system for multicellular animals) to try
out, as a random variation, the higher level of neurological organization
required for locomotor imagination. Its possibility is clearly not a foregone
conclusion. Indeed, it happened in only one kind of telesensory animal, the
vertebrates. It did not happen in invertebrate animals.
The
Inferiority of Invertebrates. The evolution of telesensory animals
in the direction of natural perfection for organisms of their kind is necessary
for the evolution of subjective animals, but it is not sufficient. The telesensory
stage must make it possible for the mechanism of embryological development
(the biological behavior guidance system for multicellular animals) to try
out, as a random variation, the higher level of neurological organization
required for locomotor imagination. Its possibility is clearly not a foregone
conclusion. Indeed, it happened in only one kind of telesensory animal, the
vertebrates. It did not happen in invertebrate animals.
It is because vertebrates seem so basically different from other animals that all the other animals are usually classified together as "invertebrates". It has long seemed that vertebrates have a greater capacity for evolution. They generally have larger and more powerful bodies, not only those that live in water, but also those on land and in the air. The closest invertebrates come to approximating the vertebrate animal system of representation are the octopus and squid, which have evolved the same kind of camera-type eyes (focusing images on a retina) that vertebrates of all kinds have.
However, many kinds of invertebrates, including insects, many mollusks, and even worms, are in the same category of natural kinds as sharks, fishes, amphibians, reptiles, birds and even dinosaurs. All are telesensory animals, with centralized nervous systems and the capacity to guide behavior in relation to objects at a distance. And map-making powers evolve in both classes of animals. Though the examples mentioned above are invertebrates, similar map-making capacities have been demonstrated in telesensory chordates, such as fish, reptiles and birds (Gallistel 1990, Chapters 4 and 5).
There is a similar difference between two classes of somatosensory animals, even those whose radial bodies indicate their adaptation at this level. There are simple somatosensory animals, which include the hydra, jellyfish, sea anemones and corals (coelenterates) and their somewhat more complex cousins, the comb jellies (ctenophora). But there is another kind of radially symmetrical animal that is far more complex, the echinoderms, including sea stars ("starfish") and sea urchins. The difference in their complexity does not come from their stage of evolution, since they are both kinds of somatosensory animals. (But it is explained, as we shall see, in the same way as the difference among telesensory animals.)
The difference in the fates of these two classes of animals poses a challenge to this ontological explanation of revolutionary evolution. If evolution is really a dialectic of gradual and revolutionary change in which stages of evolution follow one another in time, then why did the next stage not happen to any invertebrates species? Thus, let us consider what makes vertebrates so different from invertebrates before we show how the vertebrate nervous systems evolves a higher level of neurological organization.
The answer to this puzzle is found in their patterns of embryological development. There are basically two different ways that multicellular animals develop, and it has long been recognized as a basic difference among them. Once class has so-called “proterostome” development, whereas the other has “deuterostome” development. Deuterostome embryological development in vertebrates and how it differs from proterostome development will explain what makes it possible them to try out higher levels of neurological organization. The origin of this difference was explained, along with a more detailed description of the difference between proterostome and deuterostome nervous systems, in Stage 4: Origin of the mechanism of embryological development.
Since every cell in a (sexually reproducing) multicellular animal derives from the same fertilized egg cell and has the same genetic structure, the basic challenge in constructing a complex body or nervous system is to make the cells become different in their behavior and yet coordinate them so that they work together as a whole. This challenge has been met by a process called embryological development, which began with the original multicellular animals of this kind and became increasingly elaborate through evolution.
Sexual reproduction requires, as we have seen, that the whole body of the multicellular animal be constructed from a single fertilized egg cell, and this mechanism coordinates the lower level organisms of which multicellular animals are composed. This is a function of what is classified here as the "multicellular biological behavior guidance system."
The advent of that important mechanism is responsible for the Cambrian revolution, which marks the very beginning of the fossil record some 600 million years ago. That is when the fossils of many different species of multicellular animals suddenly showed up, and since fossils form only when multicellular animals evolved hard parts, like shells, it indicates roughly when the mechanism of embryological development first evolved. That is the mechanism that constructs the nervous system.
Cells are first determined to become different kinds by the part of the cytoplasm (or cortex) of the fertilized egg that they inherit from the initial divisions of the egg cell. But animals need complex bodies in order to act on other objects in space, and that requires a developmental process in which some cells (and groups of cells) move past other groups of cells that remain attached to one another. What made it possible for animals to have a form of embryological development that is so different from plants is the animal-like behavior of the protists from which animals evolved (presumably, amoebas), including locomotion. Except in the simplest animals, development involves a process of gastrulation, and early in that process, a major difference shows up among multicellular animals, indicating an early branching in the evolution of their kinds. It separates vertebrates from most kinds of invertebrates, and it explains why vertebrates are able to evolve higher levels of neurological development, beyond telesensory animals, whereas invertebrates cannot.
The major division among animals is the difference between proterostomes and deuterostomes. Most invertebrates are proterostomes, but vertebrates are deuterostomes. A brief sketch of the difference about the basic frameworks in which their nervous systems are laid out will make it clear why higher levels of neurological organization occur only in vertebrates.
In proterostomes, the basic structure produced by gastrulation is a single tube formed of two layers of cells (ectoderm and endoderm) stretching between the mouth and anus. There are apparently gradients of messenger proteins established in the tube as a whole, uniquely identifying every location in it. Although the mechanism has not yet been fully explained, the multicellular behavior guidance system can apparently address cells by their locations in this gradient, make them open up different segments of their chromosomes so they behave differently, and thereby enable them to move past other cells to other locations in the body which are also identified by this gradient. In proterostomes, this is the source of the structure of both the nervous system and most of the structure of the body. Neurons always develop only from the outer layer of cells in the tube (ectoderm). Cells located in certain parts of the two-layered tube are determined to become ganglia, which are clusters of neurons that set up their own gradients of messenger molecules among themselves. And the multicellular behavior guidance system can also use these new gradients, within the ganglia that form, to address cells individually and make them send axons to connect with certain other neurons in the same ganglion or to certain neurons in other ganglia located elsewhere in the body.
In deuterostomes, gastrulation is able to produce a basically different structure of gradients for the multicellular behavior guidance system to use in determining cells, although not all deuterostomes exploit this capacity. There is at least a 2-D gradient in the tube extending from mouth to anus identifying the locations of cells (with cells also orienting themselves orthagonally to the surface of the tube), and there is an additional body-length tube, the neural tube, with a similar 2 .D gradient of its own, which becomes the spinal chord and brain in vertebrates.
The neural tube is formed from a body-long strip of the outer layer of cells (or ectoderm) of the body-tube (or gastrula) which folds over and sinks beneath the outer layer of cells. Although neurons still develop only from the ectoderm, the ectoderm used in deuterostomes is the neural tube, and its separation from the rest of the body enables the multicellular behavior guidance system to use the 2-D gradients in the neural tube to address neurons separately from the cells that become other parts of the body.
Furthermore, during the process of gastrulation in which the neural tube is formed, both the body and the neural tube are segmented, so that there is actually a series of 2 .D gradients (including a difference between right and left sides of the body) running along the length of both the body and the neural tube. Thus, not only are all the cells of the behavior guidance system already separated from the rest of the body in a tube with their own 2 .D gradient, but that all .inclusive 2 .D gradient is also subdivided regularly into smaller 2 .D gradients. The animals that take advantage of deuterostome development to construct a nervous system in this way are all called "chordates".
Since the challenge to the mechanism of embryological development is to coordinate the behavior of lower level organisms, it is clearly an advantage to have a segmented neural tube in which to construct and operate the animal behavior guidance system separate from the rest of the body. Indeed, there is a useful analogy to the biological behavior guidance system. Chordates seem to have a higher level of organization than proterostomes in a way that resembles how eukaryotes differ from prokaryotes, although chordate and proterostome nervous systems are actually on the same level of part-whole complexity.
The prokaryote and eukaryote biological behavior guidance systems both coordinates the behavior of segments of DNA (or genes) in much the same way as the multicellular animal behavior guidance system coordinates the behavior of neurons in both proterostomes and chordates. The lower level organisms are coordinated by being parts of the structure of the organism as a whole. But prokaryotes are to eukaryotes as proterostomes are to chordates.
In prokaryotes, a loop of DNA, containing all the genes, is located alongside all the other molecules in the cell, whereas in eukaryotes, multiple chromosomes are kept separately in a nucleus, where the expression of all their genes can be coordinated without interference from the mechanisms generating the behavior of the cell as a whole.
Likewise, in the case of nervous systems. In proterostomes, neurons are determined to become ganglia from the body's 2 .D gradient alongside all the other organs of the body, whereas in chordates, the only cells that become neurons come from the 2 .D gradients of a segmented neural tube, so that the coordination of cells in setting up the nervous system is not disrupted by the coordination of cells in setting up in the body.
In both eukaryotic cells and chordates, there is a spatially distinct organ for controlling the simplest units involved in guiding behavior. It is important to recognize, however, that deuterostome development does not put chordates on a higher level of part-whole complexity than proterostomes. It does not make much sense to talk about levels of part-whole complexity when there are only two levels.
There is, moreover, a further advantage in separating the animal behavior guidance system in the neural tube from the rest of the body. In proterostomes, where there is no such separation, another set of gradients has to be set up in each ganglion to coordinate its neurons after the ganglion has been determined from the ectoderm. In chordates, by contrast, this step has already been accomplished before neurons are determined, because cells in the neural tube already contain labels indicating their relative locations in the local 2 .D gradient of each segment of neural tube as well as their segment relative to other segments in the neural tube as a whole. The chordate multicellular behavior guidance system can, therefore, address whole 2 .D arrays of cells in a segment at once and tell (some of) them (from each local area) to migrate to, or to send axons to, another 2 .D array in the neural tube, while keeping their relative positions in their own 2 .D structure and finding their counterparts in the target array. That is, chordates have 2 .D arrays located in a more inclusive 2 .D array. Thus, it is as if the units supplied by gastrulation for use in neural development were not just single neurons, but also whole 2 .D arrays of neurons.
This difference would explain why only vertebrates are able to try out yet higher levels of neurological organization, and we shall see why such connections among whole 2 .D arrays of neurons are essential to the mechanisms of the subjective animal system of representation that gives mammals a locomotor imagination. It will not be known precisely how the segmented neural tube gives vertebrates such an advantage until the mechanisms of the multicellular biological behavior guidance system are explained, but that is not essential to this argument, for it depends only on recognizing a basic difference between the biological behavior guidance systems of proterostomes and deuterostomes in how they coordinate the behavior of cells in setting up the structure of the nervous system.
The difference between a nervous system enclosed in a separate unit and one spread throughout the body is intuitively clear, and there are many reasons for believing that this accounts for the failure of invertebrates to evolve higher levels of neurological organization are not hard to find. Here are three (which are explained in more detail in Stage 4: Multicellular organisms: Evolution of embryological development).
First, it is clear that the gradients in the body of invertebrates could interfere with the structure of the nervous system, because, despite the obvious advantage to telesensory animals of having a nervous system centralized in the head for guiding locomotion, not a single proterostome has managed to disentangle its nervous system from the mouth itself. At the head of every telesensory proterostome, there are always two pairs of ganglia whose connections encircle the mouth. This reflects the gradients set up in the body by proterostome development, in which the mouth is the opening in the sphere created by gastrulation. Proterostome development is simpler than deuterostome, but it compromises the unity of the centralized nervous system.
Second, although proterostomes do develop segmentation in their bodily structures (in segmented worms and insects), it does not give the nervous system a larger 2 .D gradient in which smaller 2 .D gradients are embedded for use in connecting 2-D arrays at different locations with one another, because it is the body as a whole that is segmented. The body is, in effect, a series of simpler, proterostome bodies. Ganglia can be set up in each segment, but they still have to set up their own internal 2 .D gradients. Hence, they are independent of one another and not coordinated with one another by being embedded in a larger 2 .D gradient.
Unlike the deuterostome development of chordates, where the process of gastrulation segments both the body and the neural plate (which becomes neural tube), the segments in proterostomes are set up by the duplication of special mesodermal cells, which give rise to coeloms that divide the basic tube-like structure of their bodies.
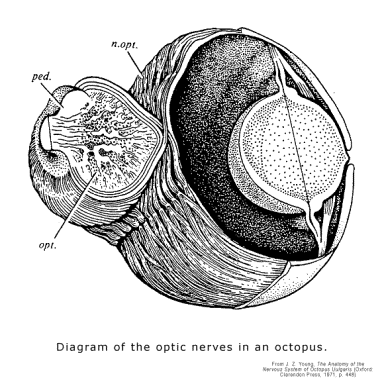
Third, the evolution of a camera-type eyes in the squid and the octopus shows that the difference between invertebrates and vertebrates cannot be explained by any lack of ability to evolve the right kinds of cells. But the difference in their wiring does show the advantage of having 2 .D arrays built into the gradients of the neural tube. In vertebrates, the retina of the eye develops from a section of the neural tube that migrates as a whole to the ectoderm (where they determine cells located there to become a lens and an eyeball surrounding it). So their axons, with millions of them in a bundle, are able to keep their spatial relations to one another as they project back to the neural tube and superimpose themselves in an orderly way on the 2 .D arrays of their targets. But in order for the octopus, as an invertebrate, to operate a camera-type eye, the complex 2 .D array of cells must be constructed from the neurons in ganglia, and whatever is going on, a glance at the basket-weave structure of its optic nerve makes clear that it takes a good part of the multicellular biological behavior guidance system's capacity for determining cells simply to keep them straight as they project to their targets. Not surprisingly, it is not easy to combine many such neural mechanisms as parts of a larger neural mechanism. (See Diagram.)
By explaining why invertebrates are unable to try out higher levels of neurological organization, we also explain why vertebrates do. Deuterostome development coordinates lower level organisms in a way that can set up a segmented neural tube separate from the rest of the body so that it can then use all its powers of coordination to set up the nervous system and only later connect it with the body (using its parallel set of segmented 2-D gradients).
Vertebrates also differ from invertebrates, of course, in having generally larger and more powerful bodies. That has not been explained. Nor will we try to explain it, except to suggest that, since their size and power are mainly a matter of having an internal skeleton, rather than an external skeleton (as in insects) or no skeleton at all (as in the octopus), and since the vertebras are also determined as a separate unit running the whole length of the body (the notochord) in the same process of gastrulation and segmentation that determines the body and the neural tube (and the coelom, from which the internal organs come), it would not be surprising if the same cause were responsible for this other difference from invertebrates as well.
![]() The
structure of the non-mammalian vertebrate brain. In the non-mammalian
vertebrate brain, the three systems serving the main subfunctions of the animal
behavior guidance system are located in three anatomically distinct parts
of vertebrate brains. It is relevant to take account of this structure, because
shifting the sensory input and behavioral output systems to the part of the
brain that served only the goal selection subfunction in non-mammalian vertebrates
is the random variation that will try out the higher level of neurological
organization found in the mammalian brain.
The
structure of the non-mammalian vertebrate brain. In the non-mammalian
vertebrate brain, the three systems serving the main subfunctions of the animal
behavior guidance system are located in three anatomically distinct parts
of vertebrate brains. It is relevant to take account of this structure, because
shifting the sensory input and behavioral output systems to the part of the
brain that served only the goal selection subfunction in non-mammalian vertebrates
is the random variation that will try out the higher level of neurological
organization found in the mammalian brain.
![]() Basic structure. In the non-mammalian brain,
the major systems and their functions are obvious, given the function of the
animal behavior guidance system (and its three subfunctions). The brain itself
is one of two main components of the nervous system set up in the neural tube.
Basic structure. In the non-mammalian brain,
the major systems and their functions are obvious, given the function of the
animal behavior guidance system (and its three subfunctions). The brain itself
is one of two main components of the nervous system set up in the neural tube.
The other component is the spinal chord. It is the entire nervous system of somatosensory chordates (represented by Amphioxus) from which vertebrates evolved. For vertebrates, the spinal chord serves mainly to connect the brain with the rest of the body.
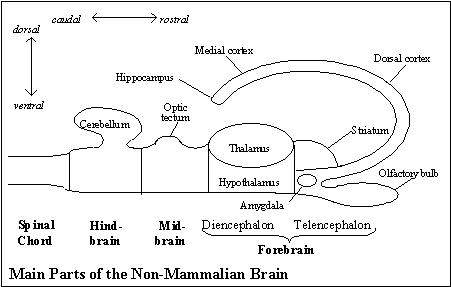
In the brain's segment of the neural tube, embryological development results in three, anatomically distinct parts: just above (rostral to) the spinal chord is the hindbrain, followed by the midbrain, and finally (at the rostral-most end) the forebrain. These three units can be explained by the three subfunctions that any system that would choose among different kinds of behavior must have. Any behavior guidance system, as we have seen, must register input from the world, select from among incompatible kinds of behavior, and generate the kind of behavior chosen. That holds for the vertebrate brain. These three subfunctions account for the kinds of neural connections found in the hindbrain, midbrain and forebrain, as a brief sketch of their connections will confirm. (See the diagram of the main parts of the non-mammalian brain.)
Furthermore, the neural connections among these sub-systems are just what would be expected of an animal behavior guidance system with an animal system of representation, that is, with input from the bodily condition being combined with telesensory input to construct an animal representation of the object that can guide locomotion in relation to distant objects.
Hindbrain’s behavioral output system. The hindbrain is responsible for motor output to the body. In order to guide locomotion by telesensory input, the hindbrain takes over control of local reflexes in all segments of the original neural tube (as in Amphioxus), and thus, the somatosensory input that triggered those local reflexes is also passed toward the head (rostrally) and registered in the hindbrain. (See diagram of the behavioral output system.)
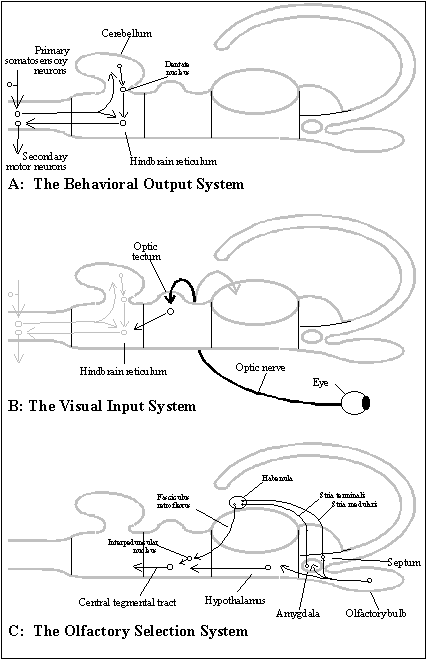
But the hindbrain does not represent the body somatotopically, that is, as a picture or isomorphic representation of the body. (Its diffuse organization is called a "reticular formation".)
There is, however, an appendage to the hindbrain, the cerebellum, whose neurons correspond roughly to the geometrical structure of the body because of how it assists in generating behavior. The somatotopic organization is required by the cerebellum's function. It modifies the hindbrain's motor output in order to even out the spatial distributions (and temporal sequences) of its ultimate commands to local muscle groups (using its own somatosensory input).
That is, when different regions of the hindbrain generating different kinds of behavior call on the same local muscle groups in the spine, the cerebellum distributes their orders spatially and temporally so that all the tasks requested are accomplished without overtaxing any local group (while keeping the body oriented in gravity, with the help of the vestibular nucleus). It has its own channel of sensory input from the body, from the inner ear about acceleration and orientation in gravity, and from nuclei (olivary nuclei in the caudal hindbrain) that appear to compare motor commands with somatosensory input. It modifies motor commands by way of its output to the hindbrain behavior generator.
Midbrain’s visual input system. The midbrain registers telesensory input from the object. The main form of telesensory input was originally vision, whose 2 .D arrays, as mentioned before, are easily set up in the neural tube. The retina is a segment of the forebrain that migrates to the ectoderm of the body and determines cells already located there to become a lens and an optic cup behind it. The retinal neurons send an ordered, 2 .D array of axons to a region of the midbrain called the "optic tectum" (and to a part of the forebrain called the "thalamus" from which the retina migrated). (See diagram of the visual input system.)
The optic tectum picks objects out from the background, a relatively easy task when either the object or the animal is moving. Its control of the direction of the eye serves the function of inquisitive behavior in the animal system of representation, though the optic tectum also receives some somatosensory input. But somatosensory input includes the "lateral line" in fishes, which can detect the pressure caused by fish swimming nearby, and there is also taste input to the optic tectum (from cranial nerves that serve the feeding apparatus), and hearing (after it evolves).
The optic tectum has a large projection to the hindbrain behavioral output system, by which the optic tectum can adjust locomotion according to the direction, motion and distance to the object in space it is tracking visually and direct routine responses to objects, as in feeding. In non-mammalian vertebrates, in short, the optic tectum serves as the animal representation, or the input subfunction in the animal system of representation, for it guides the locomotion of the body in relation to the currently perceived object.
Forebrain’s olfactory selection system. The basic need responsible for the evolution of the behavior guidance system is choosing the goal, and telesensory animals must choose which goal to pursue with respect to the object at a distance (feeding, self-protection, mating, and the like). This subfunction is served in non-mammalian vertebrates by the forebrain.
In the primitive vertebrate brain, these decisions depend mainly on the forebrain's own form of telesensory input, olfaction. The olfactory bulb, located at the extreme rostral end of the telencephalon, contains neurons that identify molecules by interacting with them directly. About organisms, olfaction is usually a reliable indicator of the kind being encountered. The olfactory bulb has multiple pathways by which it reaches the midbrain and can control the kind of behavior that the hindbrain generates in response to the visual input from the object. (See diagram of the olfactory selection system.)
One pathway by which the olfactory bulb affects the midbrain is through the hypothalamus, a (ventral) part of the diencephalon, which has multiple connections to the midbrain. A second pathway is through the amygdala, another nucleus in the telencephalon, which, as the seat of fear, can override other routines and generate fighting or fleeing instead. And there is a third pathway involving the septum, a ventral part of the telencephalon.
In primitive vertebrates, the main route for affecting the midbrain is via the habenulae, a prominent, asymmetric pair of nuclei located just above the diencephalon (or atop the dorsal thalamus) which has a heavily myelinated bundle of fibers (the fasciculus retroflexus) that passes through the thalamus to a spiral shaped nucleus at the ventral midline of the midbrain (the interpeduncular nucleus). The olfactory bulb projects to the hypothalamus and the septum (which are interconnected), and they have connections to the habenulae via the stria medularis. The amygdala (which is also connected to the hypothalamus) has a connection of its own to the habenulae via the stria terminalis. (See diagram of the olfactory selection system.)
The forebrain also has, as we shall see, two other ways of affecting motor output, which depend on other kinds of sensory input. The forebrain receives (via its thalamus) visual input (both indirectly, by way of the optic tectum, and directly, from the retina), auditory input (after hearing evolves), and even some somatosensory input, all of which become increasingly pronounced in the evolution of vertebrates. (See diagrams of the forebrain output.) But these sensory inputs are used only to modify behavior in relation to distant objects currently being detected by the optic tectum and to generate complex instinctive routines, including telesensory map-making routines. Thus, there are, including olfaction, three ways that the forebrain can influence what happens in the midbrain and hindbrain, and all have to do with the subfunction of selecting the right kind of behavior.
![]() Foundation
for a higher level of neurological organization. Looking ahead, the
transformation of the non-mammalian into the mammalian brain is often described
as the centralization of functions in the nervous system. That is, as mentioned
above, the three subfunctions of the animal behavior guidance system, which
are parceled out to the three anatomically distinct sections of the non-mammalian
brain, are all served by one of those sections in mammalian brains, namely,
by the forebrain. The midbrain and hindbrain structures are still there, but
they are slaves to the forebrain. The simplest and most convincing way to
show how the four subsystems of the subjective animal system of representation
are the functions served by structures in the mammalian forebrain will be
to describe how those structures evolve from simpler mechanisms in the non-mammalian
forebrain, and thus, it will be helpful to describe the relevant structures
of the vertebrate brain here.
Foundation
for a higher level of neurological organization. Looking ahead, the
transformation of the non-mammalian into the mammalian brain is often described
as the centralization of functions in the nervous system. That is, as mentioned
above, the three subfunctions of the animal behavior guidance system, which
are parceled out to the three anatomically distinct sections of the non-mammalian
brain, are all served by one of those sections in mammalian brains, namely,
by the forebrain. The midbrain and hindbrain structures are still there, but
they are slaves to the forebrain. The simplest and most convincing way to
show how the four subsystems of the subjective animal system of representation
are the functions served by structures in the mammalian forebrain will be
to describe how those structures evolve from simpler mechanisms in the non-mammalian
forebrain, and thus, it will be helpful to describe the relevant structures
of the vertebrate brain here.
There are two important kinds of changes in the mammalian brain, not only the evolution of a neocortex, but also the redirection of erstwhile forebrain output back to the forebrain (by way of the thalamus) to form the complete circuits of its higher level of neurological organization. To see what is involved in these changes, let us look at the structure of the non-mammalian forebrain more closely and its three outputs to the midbrain.
Embryological development of the forebrain. The vertebrate forebrain develops from the extreme rostral section of the neural tube. It has two parts, the diencephalon and telencephalon. (See earlier diagram of the main parts of the non-mammalian brain.)
The diencephalon is the neural tube itself at that most rostral section. It is divided into two parts: the dorsal half, the thalamus, receives and passes on to the telencephalon all the forebrain's sensory input, and the ventral half, the hypothalamus, is the master control of the endocrine system (managing the messenger proteins, or hormones, in the blood) and the parasympathetic nervous system (sending nerves to internal organs). (The hypothalamus is, therefore, the main way in which the outputs of the animal behavior guidance system and the biological behavior guidance system are coordinated.)
The telencephalon develops from the same segment of the neural tube as the diencephalon. That segment of the neural plate does not close fully into a tube like the rest of the nervous system; instead, both sides are extended and thickened and become two, symmetrical flaps, which ultimately almost surround the diencephalon, as the telencephalon.
Forebrain output to the midbrain. The non-mammalian forebrain projects all its output to the midbrain, because that is where the optic tectum uses telesensory input to guide the locomotion in relation to the distant object (that is, where the animal representation of the object guides locomotion). The default behavior is normal locomotion and feeding. (Taste receptors have direct connections that affect feeding behavior.) Output from the forebrain modifies these behaviors and is responsible for selecting most other goals. The forebrain's outputs are all received by a region of the midbrain where the optic tectum influences the hindbrain motor commands and determines how the body moves in relation to the distant objects, approaching or avoiding them.
The most primitive output of the non-mammalian forebrain to the midbrain for choosing the goals of the behavior has already been mentioned. It originates from input to the olfactory bulb and reaches the midbrain by various ancient pathways, including some through the hypothalamus. This, together with another olfactory pathway (through the amygdala, a ventral part of the telencephalon), becomes the affective system in mammals.
It is the two other ways that non-mammalian forebrains can influence behavior that are relevant to the evolution of the subjective system of representation itself. Both of these other two outputs depend on visual, auditory or somatosensory input received by the thalamus and relayed to the telencephalon. But each involves a different target in the forebrain. One target is the (non-mammalian) dorsal cortex, the extended flap that develops from the neural tube at the diencephalon into the telencephalon, and the other is the striatum, a structure at the base of that extended flap. The dorsal cortex sends output to the midbrain by way of its connections to the extreme (medial) edge of the flap, which is (or becomes) the hippocampus. And the hippocampus projects, in turn, (by way of the fornix) to the hypothalamus, which sends output to the midbrain and to the mammillary body located in the hypothalamus. (The mammillary body has its own projection to the midbrain.) (See diagram of the hippocampal and striatal two pathway for forebrain output.)
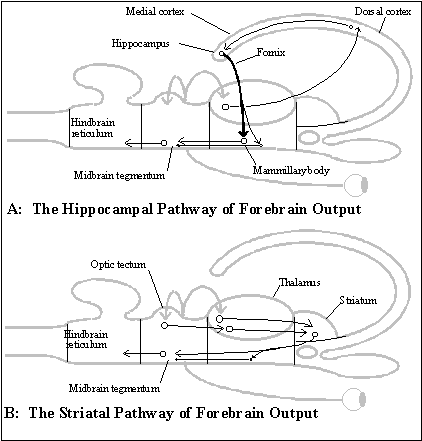
The striatum, the other forebrain target, sends output to the midbrain by way of connections in and through the hypothalamus. (See diagram of the hippocampal and striatal two pathway for forebrain output.)
The size of the forebrain increases in vertebrate evolution, and the size and definition of the striatal and hippocampal pathways to the midbrain increase more than that of the primitive olfactory pathways. But the function of striatal and hippocampal output to the midbrain is basically to select the right kind of behavior for the distant object, including the construction of instinctive telesensory maps. They generate instinctive responses, including complex fixed action patterns, by modifying behavior that is already being adjusted to the currently detected distant object by the optic tectum in the midbrain.
For example, in an amphibian like the frog, visual input from the retina and optic tectum that is relayed (by the thalamus) to the striatum is used to select flight from large objects that are moving toward them (which are usually predators), suppressing their normal behavior of snapping at food. And visual input from the retina relayed (by the thalamus and dorsal cortex) to the hippocampus is used to keep frogs from jumping into walls. (See Macphail 1982 on amphibians and reptiles.)
In birds, at least, this hippocampal pathway evolved into a complete circuit for a one-dimensional, vision-based memory for retracing a pathway of salient landmarks, which approximates the mammalian memory circuit.
In the avian brain, the dorsal cortex contains a detailed visual image called the "visual Wulst" which controls locomotion by its own, direct motor output. It is part of a memory map that uses visual input relayed by the thalamus directly from the retina to the dorsal cortex. And as part of the dorsal cortex, the Wulst connects through the medial cortex/hippocampus and fornix back to the mammillary body in the hypothalamus. But instead of merely influencing the midbrain and motor output, as in other non-mammals, the mammillary body relays a signal, by way of the thalamus, back to the Wulst in the telencephalon. This is a complete circuit, resembling the mammalian memory mechanism, is apparently used to construct a memory map for guiding locomotion. Assuming it works the same way as in mammals, the Wulst can use a one-dimensional chain of stimulus-response connections as a map to guide locomotion without involving the rest of the circuit (hippocampus, fornix and mammillary body). But vision is the only sensory modality involved, and judging by the neurological mechanisms, it cannot use its memory map as a locomotor imagination.
Reptilian innovations. There is little change in the basic structure of the non-mammalian vertebrate brain (after the evolution of the jaw in fishes), except for three important changes, each involving one main parts of the brain, that came with the evolution of reptiles and the their use of legs to support the body.
Legs that could support the body made vertebrates better able to move across land, and new mechanisms were required to operate them. A new section of the cerebellum (the paleocerebellum) was required to balance the body on its legs and smooth out motor commands to them. And to serve this function, it has a somatotopic organization, that is, like a picture of the body.
The paleocerebellum projects to a new nucleus in the midbrain, the red nucleus, which uses a somatotopic array of neurons to send motor commands to the limbs.
Finally, the source of the more complex motor commands required to operate legs is a prominent, new structure in the telencephalon, the DVR (dorsal ventricular ridge). It is a major modification of the striatal pathway of forebrain output, because the DVR receives most of the thalamic relay of sensory input via this pathway to the telencephalon (including both the direct and the indirect visual channels), and the DVR projects to the striatum, by way of which its output reaches the midbrain and hindbrain areas for generating motor output. The DVR controls the motor commands from the red nucleus by way of a projection from the hindbrain to the paleocerebellum. (The main part of the DVR also projects to a more basal part of the DVR, which also receives olfactory input and projects to the hypothalamus, according to Ulinski, 1983.) (See diagram of the reptilian brain with paleocerebellum, red nucleus and DVR.)
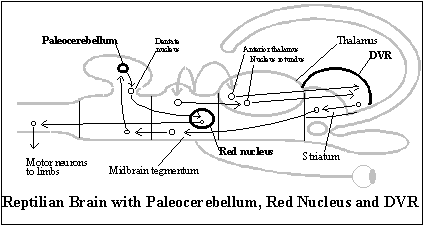
To sum up, there are only three ways that the non-mammalian forebrain can affect the behavior being generated in relation to the distant object by the midbrain and hindbrain: the ancient olfactory pathway, the hippocampal pathway, and the striatal pathway. And the only input to the non-mammalian forebrain is the sensory input (indirectly from the midbrain and directly from the retina) registered in the thalamus of its diencephalon and relayed to various structures in the telencephalon — that is, except for the forebrain's own telesensory input, olfaction, and what the hypothalamus gathers from its control of the endocrine and parasympathetic nervous system. Although there is, as we have seen, a further development of the reptilian brain in birds, this is the basic structure of the reptilian forebrain from which the subjective animal system of representation evolves.
![]() The
gradual evolution of vertebrates. In the first section of this
chapter, we found that the evolution of telesensory from somatosensory animals
is inevitable because it is both possible and functional. In the second section,
we saw how deuterostome development of chordates makes it easier to construct
a nervous system with a telesensory level of part-whole complexity, and the
third section described in detail the structures in the vertebrate brain that
are the foundation for using the deuterostome capacity to determine entire
2-D arrays of neurons to construct a brain with yet higher level of part-whole
complexity. We are following the inevitable stages of evolution that lead
up to beings like us, and thus, it remains only to trace the gradual evolution
of telesensory animals on earth. We may set aside animals with proterostome
embryological development, because we have sufficient reason to believe that
they are not capable of evolving beyond the telesensory stage. Let us consider
briefly, therefore, the origin and evolutionary career of vertebrates, beginning
with the somatosensory animals from which they evolved.
The
gradual evolution of vertebrates. In the first section of this
chapter, we found that the evolution of telesensory from somatosensory animals
is inevitable because it is both possible and functional. In the second section,
we saw how deuterostome development of chordates makes it easier to construct
a nervous system with a telesensory level of part-whole complexity, and the
third section described in detail the structures in the vertebrate brain that
are the foundation for using the deuterostome capacity to determine entire
2-D arrays of neurons to construct a brain with yet higher level of part-whole
complexity. We are following the inevitable stages of evolution that lead
up to beings like us, and thus, it remains only to trace the gradual evolution
of telesensory animals on earth. We may set aside animals with proterostome
embryological development, because we have sufficient reason to believe that
they are not capable of evolving beyond the telesensory stage. Let us consider
briefly, therefore, the origin and evolutionary career of vertebrates, beginning
with the somatosensory animals from which they evolved.
![]() Somatosensory
chordates. We are counting somatosensory animals as a necessary part
of the multicellular biological stage of evolution, because we want to count
how many distinct levels of part-whole complexity and, thus, how many distinct
evolutionary stages, there are in the ladder that begins with the original
proto-organisms and ascend to beings like us. The multicellular animals from
which vertebrates evolved are somatosensory chordates, and they are represented
by the brainless chordate, Amphioxus.
Somatosensory
chordates. We are counting somatosensory animals as a necessary part
of the multicellular biological stage of evolution, because we want to count
how many distinct levels of part-whole complexity and, thus, how many distinct
evolutionary stages, there are in the ladder that begins with the original
proto-organisms and ascend to beings like us. The multicellular animals from
which vertebrates evolved are somatosensory chordates, and they are represented
by the brainless chordate, Amphioxus.
Without a brain, its nervous system is equivalent to the vertebrate's spinal chord. Relatively sophisticated swimming behavior is generated in its bilateral body by local connections between segments of its neural tube, each with sensory, motor and interneurons serving the three basic subfunctions of somatosensory animal behavior guidance systems. The function of selecting the right kind of behavior is served by a cough reflex at the mouth that rejects unwanted particles. And motor reflexes triggered by somatosensory input lead Amphioxus to burrow into sand at the edges oceans around the world in order to feed on microorganisms and protect itself.
Before vertebrates evolved, brainless chordates must have acquired free energy in more active ways than Amphioxus, feeding on microorganisms while swimming or seeking out multicellular plants to browse on. It is not known how vertebrates evolved from animals like Amphioxus, except that it was part of the evolution of telesensory organs. But it is no surprise that Amphioxus is the only extant representative of the first stage, because when chordates did evolve telesensory organs and a centralized nervous system to use them in guiding locomotion, somatosensory chordates were left with only the most low-energy, sedentary ecological niches.
![]() Telesensory
chordates (Vertebrates). According to the detailed fossil record, which
begins with the Cambrian Period about 570 million years ago, vertebrates showed
up about 500 million years ago, after almost every kind of marine invertebrate
had already evolved.
Telesensory
chordates (Vertebrates). According to the detailed fossil record, which
begins with the Cambrian Period about 570 million years ago, vertebrates showed
up about 500 million years ago, after almost every kind of marine invertebrate
had already evolved.
Multicellular plants had evolved long before the Cambrian period, and what accounts for the fossils of animal skeleton that mark the beginning of that period is the evolution of the mechanism of embryological development (that is, the biological behavior guidance system for multicellular animals). The relatively late arrival of chordates suggests that deuterostome development evolved after proterostome development.
Vertebrates are named after the series of vertebra that enclose their neural tube, except at the head, where the brain is located. From the beginning, as we have seen, the vertebrate brain had three, anatomically distinct parts (midbrain, forebrain and hindbrain), serving, respectively, the three subfunctions of the animal behavior guidance system (input, choice and output), and we have seen how the function of the telesensory animal system of representation is served by its structure. There is remarkably little change in the structure of the non-mammalian brain, but the bodies of non-mammalian vertebrates underwent radical changes.
The most primitive vertebrates, jawless fishes (Agnathia), were still filter feeders. They actively scooped up detritus from the bottom, although their contemporary descendants, lampreys, have adapted the jawless mouth for preying on other fish.
From them evolved the now extinct, heavily armored placodermi, with a major reorganization of the hindbrain to operate its hinged jaw. The jaw not only made them ferocious predators, but also revolutionized the means of ingesting other organisms for all subsequent vertebrates.
Two kinds of fishes evolved from placodermi, cartilaginous (such as, sharks and rays) and bony. The cartilaginous skeleton was a departure from the bony frames of placodermi and jawless fishes. But bony fishes were so successful that there may now be as many as 40,000 species of them. The earliest of them were fresh water fishes, which had lungs as well as gills to supplement their supply of oxygen when the water was stagnant. In most extant species, the lungs have evolved into a swim bladder, which is also used for water-borne hearing. But one early lung fishes, the lobe-fin fishes (Crossopterygii), had fleshy bases for their two pairs of fins.
From lung fishes evolved amphibians. Their ability to pull themselves across the land opened up a new range of living objects to ingest, for by this time, plants and some insects had already invaded the land. (They were our first example of gradual evolution by reproductive causation, in Change: As aspect of substance: Global regularites: Reproductive: Gradual evolution).
There was room for improvement among vertebrates that completed cycles of reproduction on land. Although fertilization is still an external, water-based process in amphibians, it was done internally by reptiles. Reptiles not only had eggs with a shell that protected them from drying out as the embryos developed on land, but the reptilian body also acquired a dry, scaly, relatively impermeable skin. There were also advances in the heart and lungs. And whereas amphibians had short, weak limbs attached high on the sides of their bodies, reptiles evolved longer and stronger limbs that push directly downward, enabling their bodies to clear the land as they moved along. In order to operate these limbs, new structures evolved within the basic three-part structure of the vertebrate brain to send different motor commands to different legs (the paleocerebellum in the hindbrain, the red nucleus in the midbrain, and the DVR in the forebrain) . Reptiles also have air-born hearing, which among amphibians, occurs only in frogs.
Dinosaurs and birds evolved from reptiles. Dinosaurs were apparently able to move around very quickly on two legs, freeing their front legs to grasp other living objects; only as larger dinosaur species evolved did some return to four-footed locomotion. Dinosaurs may have evolved some control over bodily temperature, but we have little evidence apart from birds, which descend from an early dinosaur species. Hollow bones and feathers adapted birds to flight, and besides improvements in heart and lungs, they acquired the metabolism to maintain a high body temperature. Since hatchlings cannot acquire energy for themselves until their bodies have developed far enough to fly, birds also care for their young, a rather rare trait before the evolution of mammals. Birds also have, as we have mentioned, a one-dimensional, stimulus-response, vision-based memory for retracing a pathway, as in returning to a nest, patrolling a territory, or migrating with the seasons, and dinosaurs probably did too.
Although these changes in vertebrates required enormous periods of time, they are all at the same stage of animal evolution. Jawless fishes first appeared about 500 million years ago, about 70 million years after the start of the Cambrian period. The jaw evolved somewhat more than 400 million years ago, about the time that plants invaded the land. Amphibians evolved from the lobe-fined lung fish a little more than 350 million years ago, during the age of fishes. Reptiles evolved about 300 million years ago, late in the age of amphibians, and the dinosaurs evolved about 225 million years ago, after the decline of amphibians, followed shortly by birds (over 200 million years ago). The evolution from fishes to amphibians to reptiles to dinosaurs to birds is not a result of the dialectic of gradual and revolutionary change, because their nervous systems (and bodies) are on the same level of part-whole complexity. They are just a series of adaptations of an animal behavior guidance system with a telesensory animal system of representation (and the body it operates) to new habitats on land and in the air (though some used their improved design to return to water). These changes did not alter radically the structures of their brains.
![]()