![]()
 Neurological
Stages of Evolution. We have used the series of levels of biological
organization to trace the course of evolution up to the stage of multicellular
animals, and, together with the explanation of the animal’s need for an additional
behavior guidance system, we have catalogued all the basic kinds of animals
that come to exist in the course of evolution, including the seemingly anomalous
animals. At this stage of biological evolution, we take up the evolutionary
stages that depend on levels of neurological organization, and as we have
seen. Those stages fall into two distinct series, the animal stages of
evolution and the spiritual (animal) stages of evolution, depending
on whether they also involve the evolution of an animal on the next higher
biological level as well. The animal stages of evolution will be discussed
here, and the new wrinkle involved spiritual stages of evolution will not
be discussed further until we take up the eighth stage, primitive spiritual
animals.
Neurological
Stages of Evolution. We have used the series of levels of biological
organization to trace the course of evolution up to the stage of multicellular
animals, and, together with the explanation of the animal’s need for an additional
behavior guidance system, we have catalogued all the basic kinds of animals
that come to exist in the course of evolution, including the seemingly anomalous
animals. At this stage of biological evolution, we take up the evolutionary
stages that depend on levels of neurological organization, and as we have
seen. Those stages fall into two distinct series, the animal stages of
evolution and the spiritual (animal) stages of evolution, depending
on whether they also involve the evolution of an animal on the next higher
biological level as well. The animal stages of evolution will be discussed
here, and the new wrinkle involved spiritual stages of evolution will not
be discussed further until we take up the eighth stage, primitive spiritual
animals.
The ontological cause of the animal stages of evolution is, as we have noted, less basic than the ontological cause of the stages of biological evolution, because the causes are levels of part-whole complexity that occur within the animal behavior guidance system at the multicellular level, that is, in the nervous system.
Each new stage is caused by a higher level of part-whole complexity in the organization of neurons, and though that means that what evolves is a basically different kind of animal behavior guidance system, new animal behavior guidance systems are quite different from new biological behavior guidance systems, because they do not have to coordinate the behavior of the lower level material structures of which they are composed. What enables the biological behavior guidance system to guide the behavior of a higher level organism as a whole is coordinating the behavior of the lower level organisms of which it is part. (And as we have seen, they always use the reproductive behavior of their parts in order to generate some of its non-reproductive behavior). These animal behavior guidance systems are composed of lower level organisms, namely, neurons, which are organisms at the eukaryotic level. But their behavior (including their capacity to reproduce) has already been coordinated by the multicellular biological behavior guidance system in constructing the nervous system during embryological development. (And higher level structures of neurons are not even able to reproduce as a whole, that is, apart from the multicellular animals which they are part.)
Instead of coordinating the behavior of neurons, animal behavior guidance systems use the structure that neurons have been given in embryological development, for it is the structure of the nervous system that enables them to guide the behavior of the animal in acting on other objects in space and moving around in space. Neurons can be used to guide behavior, because they have another kind of behavior, called “firing,” by which they can affect one another. Neurons are structural causes of an action potential that propagates along their long axons and that can, upon arriving at a synapse, cause other neurons to fire or muscle cells to contract. (That is, the connection between the stimulation that makes the neuron fire and the neuron’s effects on other cells is an irreversible structural global regularity, or a disposition. and the neuron’s structure is the ontological cause that explains the connection between the efficient cause and its effect.) But what neurons do to other cells when they fire depends on how they are laid out in the structure of the multicellular body, that is, which other cells they affect, and that structure is a contribution of the biological behavior guidance system.
It is levels of part-whole complexity in the structure of the nervous system that cause the series of stages in the evolution of multicellular animals. It is a minor series by comparison with the stages of biological evolution, for it contained within one of the biological stages. But these animal stages are no less essential to the overall course of evolution in the direction of the natural perfection of life. And as in the case of stages of biological evolution, in order to show that animal stages are inevitable, it is necessary to show that each higher level is both possible and functional.
 Animal
systems of representation. The challenge of showing the inevitability
of animal stages of evolution is just opposite to the challenge in the case
of biological stages. It is the function, rather than the possibility of such
levels that is problematic.
Animal
systems of representation. The challenge of showing the inevitability
of animal stages of evolution is just opposite to the challenge in the case
of biological stages. It is the function, rather than the possibility of such
levels that is problematic.
The possibility of another level of biological organization was the main challenge in showing the inevitability of another stage of biological evolution. But the possibility of higher levels of neurological organization is not very problematic (though there is still something to be shown). The nervous system is constructed by the multicellular biological behavior guidance system, and since that is the mechanism of embryological development, even very complex nervous systems do not seem to be beyond its reach. (And as we shall see, the inevitable limits to the complexity of multicellular structures that can be constructed by it are overcome by a form of reproductive causation that is contained in the nervous system as parts of the process of neurological development.)
On the other hand, whereas the function of higher levels of biological organization was obvious in the size and scale of the behavior they afford, the function of levels of higher levels of neurological organization is not obvious. Indeed, most levels of neurological organization are not even currently recognized in neurophysiology. We begin, therefore, with the basic function of higher levels of neurological organization.
The basic function of all higher levels of neurological organization is to serve as an animal system of representation. This function derives from the nature of animal behavior, and thus, it is part of the animal behavior guidance system. And more complete ways of serving this function depend on higher levels of neurological organization.
Animals need an animal behavior guidance system, because in order to acquire the free energy needed to fuel their reproductive cycles, they must act on other object in space. They cannot sit back and absorb photons like plants. But in order to guide behavior to act on other objects in space, the animal behavior guidance system needs to take into account not only the kind of object, but also its location. Though the kind of object is relevant in selecting the kind of behavior to generate, its location is relevant in generating behavior that is directed at the object, whatever goal is chosen. And this spatial aspect of its behavior is, as we have seen, a source of potentially great animal power.
Increased animal power can come from making the spatial aspects of their behavior depend on the spatial aspects of the situation with which it will engage. By imposing a geometrical structure on a wide region of space, such animals can use the free energy there to do work — much like a giant machine in the region, except by the more efficient means of moving an animal body around in space and acting on particular objects locally.
In the first instance, therefore, what the animal needs to be more powerful is some way of determining how to move around in space relative to objects in order to put its body in a position to act in the appropriate way, whatever its goal. Even more power to cause changes in a large region that would otherwise not occur would come from being able to anticipate the effects of motion on locations. Still more power would come from the ability to anticipate the effects of the geometrical structures of objects on how they interact. Though such spatially adapted animal behavior would require more free energy than sedentary life, the additional relevant conditions it could control more than covers the cost.
The mechanism that actualizes this possible power must be part of the system that is responsible for guiding animal behavior, that is, part of the animal behavior guidance system.
Like any behavior guidance system, as we have seen, the animal behavior guidance system must serve three subfunctions in order to select between incompatible kinds of behavior. It must register input, select the appropriate kind of behavior, and generate output. In multicellular animals, these subfunctions are served by systems made of neurons, and they will be called, respectively, the sensory input system, goal selection system, and behavioral output system.
Sensory input already has the function of representing the object in the animal behavior guidance system. But in order for animal behavior to be adapted to the spatial aspects of the situation, the sensory input system must work together with the behavioral output system, as a system for representing all the possible ways adapting animal behavior spatially.
Sensory input must enable the animal behavior guidance system to select the appropriate kind of behavior toward the objects that the animal encounters. The object may be food, a predator, a mate, or whatever, and the animal behavior guidance system must generate behavior that is appropriate to it, such as, ingesting the object, escaping it, mating with it, or whatever. (Though animal behavior includes mating, it is all non-reproductive work as far as the biological behavior guidance system is concerned, because animal behavior, as such, is just a special kind of behavior generated during the growth phase of the reproductive cycle. Mating is just another way of preparing the conditions for the biological behavior of sexual reproduction.)
But whatever the goal of animal behavior, animal behavior is more powerful when it is adapted to spatial aspects of the situation, and that is another function of sensory input. But to serve this function, the sensory input system must work together with the behavioral output system, that is, as an animal system of representation.
“Animal” system of representation is an appropriate name for this mechanism, since its function is to represent the objects on which animals must act so that their behavior can be guided spatially in relation to it. Since the object is represented by a system that is based on an interaction between the sensory input and behavioral output subsystems of the animal behavior guidance system, the animal system of representation is more than just a picture of the object. It gives the animal a conception of the object. The object is what its behavior is directed at, but if we set aside the specific goals it may have in regard to the object, it is represented in such a way that behavior can be guided in relation to it. And the conception is more complete, the more behavior is adapted to spatial aspects of the object, which depends on the animal’s level of neurological organization.
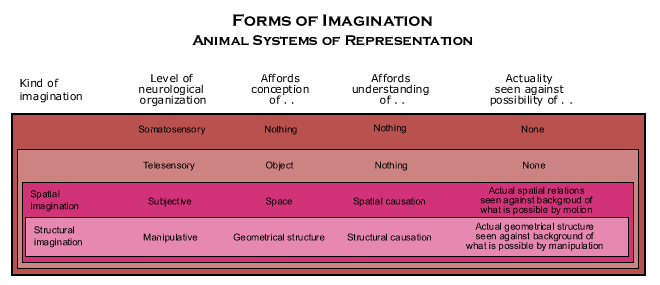
The simplest animal system of representation occurs in the “telesensory animal,” that is, animals that can perceive objects at a distance from their bodies. But the telesensory animal’s conception of the object is not a conception of the object as located in space, even though the object is typically located at a distance from the animal’s body and it guides locomotion in relation to the object. Though the object itself must be represented explicitly in order to determine its kind, its location is represented only implicitly in the instinctive routines by which the animal representation guides locomotion in relation to it.
The ability to think of the object as being located in space requires a conception of space. That is provided, as we shall see, by a faculty spatial imagination which enables the animal to think about the effects of locomotion (and motion generally) on the relations among objects in space. That occurs in subjective animals, and it is what I will call a conception of the object as located in space, because it enables them to see the object against the background of what is possible as a result of objects moving in space. (That is, spatial imagination uses sequences of images over time to represent the consequences of locomotion (and turning), and such sequences are the background in which objects are represented.) However, though the object may have a geometrical structure, the subjective animal does not have a conception of the object as having a geometrical structure. It has only the conception of the object as located in three dimensional space.
The ability to think about the object as having a geometrical structure requires a conception of geometrical structure. That is provided, as we shall see, by a faculty of structural imagination which enables the animal to think about the effects of manipulating the object on how it appears, how its structure changes, and how it relates to the geometrical structures of other objects in space. The actual geometrical structure is seen against the background of what is possible by manipulation (once again, by using sequences of images over time to represent the kinds of events that manipulation would cause). This new faculty occurs in what will be called manipulative animals, and since they already have a faculty of spatial imagination, it gives them a conception of the object in space as having a geometrical structure, for it can see the object against the background of what its geometrical structure makes possible.
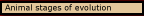 Animal
stages of evolution. Each way of these representations of the object
serves the function of an animal system of representation more completely,
but each depends on a higher level of neurological organization. These levels
are the ontological cause of the animal stages of evolution, and each requires
certain additional subsystems to serve their increasingly demanding functions.
I will use their functions to introduce the animal stages of evolution. And
the nature of somatosensory animals will be explained by contrast to all of
them.
Animal
stages of evolution. Each way of these representations of the object
serves the function of an animal system of representation more completely,
but each depends on a higher level of neurological organization. These levels
are the ontological cause of the animal stages of evolution, and each requires
certain additional subsystems to serve their increasingly demanding functions.
I will use their functions to introduce the animal stages of evolution. And
the nature of somatosensory animals will be explained by contrast to all of
them.
Somatosensory animals. Before considering what is required to serve the function of the animal system of representation, it should be emphasized that it is possible for animals not to have an animal system of representation. That is, even the object (not just its location) may be represented only implicitly in the animal behavior guidance system. In somatosensory animals, with the simplest animal behavior guidance systems, sensory input is still caused by the object, and, as always, sensory input is used to select the goal of animal behavior. But sensory input does not represent its cause as an object. At most, it might be said to represent the goal of the behavior it triggers, since it can select the kind of behavior. What serves the function of the system for representing the object is, in effect, the structure of the animal’s body.
In the somatosensory animal, it is contact with the object that guides animal behavior. In its decentralized nervous system, the object can be represented by a single somatosensory neuron, because its behavior is selected and generated by local connections between somatosensory input and motor neurons controlling nearby parts of the body. When contact reveals that the object is of the appropriate kind, behavior acts on the object at the location from which the input about its kind comes. It is a local reflex, though it may recruit appropriate behavior in other parts of the body. And reciprocally, similar reflexes in other parts of the body may trigger its motor neurons. But no centralized mechanism for guiding the behavior of the whole animal is needed. It is just what results from an equilibrium among many localized systems for guiding animal behavior.
Somatosensory animals include hydra, jellyfish, sea stars and brainless chordates (like Amphioxus). When an edible object brushes up against one of the hydra's tentacles, for example, it responds by stinging them and contracting. That reflex recruits other tentacles to contract as well, and thus, together, they pull the object contacted into its gastrovascular cavity. Sea stars also have locomotion, but it is guided locally, as each arm recruits the other arms (and its many little feet) to move in a certain direction because of the kinds of objects it contacts.
With only an implicit representation of the object, the somatosensory animal does not have a conception of the object. Though it responds according to the kind of object, no nervous mechanisms register input about its location in order to guide behavior in relation to it, because the object is always located where the sensory input is received and behavioral output is generated. Even less do somatosensory animals have a conception of the object as located in space, for it is the object’s location in space that causes it to be represented as an object at all.
Since the object need only be at a certain location to be represented as an object at all, it is the biological behavior guidance system that gives the animal its merely implicit representation of the object. Not surprisingly, therefore, this is also how behavior is “guided” to act on objects in animals at lower levels of biological organization, such as animal-like protists and bacteria. In paramecia, for example, the system for selecting whether to ingest objects or not is located in the oral groove where the object comes into contact with the animal. And the amoeba engulfs food particles wherever on its body they are encountered.
The most basic way that an animal system of representation makes animals more powerful than somatosensory animals is the ability to guide locomotion in relation to distant objects.
Though somatosensory animals may have locomotion, it is not guided in relation to distant objects in space, and thus, the geometrical structure they impose on the thermodynamic flow in the region is still short-range, channeling only the free energy of objects they happen to contact to fuel their reproductive cycles and avoiding only local hazards.
But when locomotion is guided in relation to distant object, what imposes a geometrical structure on the thermodynamics flow in the region is animal behavior itself, rather than merely the structure of the animal body. But guiding locomotion requires having a representation of the object that indicates its location relative to the body even when it is at some distance from the animals.
Sensory input from the distant object is obviously required for an animal representation of it. But in order to be useful in guiding locomotion, sensory input must be able to reveal its kind as well as its location. Unless the animal can also tell what kind of behavior to generate in relation to the object, guiding locomotion in relation to it will not control relevant conditions. In what follows, I will assume that the animal representation also indicates the kinds of distant objects as, not only their locations.
Telesensory animals. Telesensory animals are the simplest animals with an animal system of representation. The most obvious indication of their higher level of neurological organization is that their nervous system is centralized, whereas somatosensory animals have decentralized nervous systems. Centralization is required in order to guide behavior in relation to objects located at a distance from their bodies, because only groups of motor neurons can generate whole body behavior, such as swimming in fish or walking in land animals, and the same sensory input must control all of them. Sensory input also requires a higher level of neurological organization, because only groups of sensory neurons, as in the retina of the eye, can register effects of distant objects in a way that reveals information about their kinds and locations.
Sensory input from distant objects does not, by itself, contain enough information to guide locomotion in relation to it. In order to determine the object’s location, there must be an interaction between mechanisms serving the subfunctions of sensory input and behavioral output in the animal behavior guidance system. (Their interaction is also where we will find the higher level of neurological organization that gives animals spatial imagination, and the yet higher level that gives animals structural imagination.)
Sensory input from an object located at a distance from the animal, or “telesensory input,” can guide locomotion only when it is combined with input about the current condition of the animal’s body. For example, to use visual input to determine what is located in different directions, animals with eyes must take account of which way their heads or eyes are pointing when visual input is received, and that requires input about how the current condition of their eyes, head and body. I will call it input about the “bodily condition.” Thus, the animal representation of the object is telesensory input registered in a way that indicates the bodily condition. That is the animal’s “perception” of the object.
Since the animal must behave in various ways to gather information about what is located in each direction, perception generally involves a new kind of behavior. This holds of hearing and olfaction as well as vision. I will call it “inquisitive behavior,” in order to distinguish it from behavior directed at more basic goals, such as acquiring food, protection, mating and the like. Inquisitive behavior transforms telesensory input into feedback from its own motor output, making perception an active process. But whenever telesensory input from the object is registered in the nervous system according to the bodily condition at the time, it contains information that can be used to guide locomotion in attaining its goals with the object. This combination is an animal representation, because the animal can extract information from it about which way to move its body in order to change its spatial relation to the object appropriately. It is a perception of the object that implicitly contains information about its location.
Animal representations of objects also make it possible for telesensory input from an object at one time (when the object has one location relative to the animal), to be used in guiding the animal's behavior at another time (when the object has a different location relative to the animal). And in some telesensory animals, such as bees, so much information about the locations of objects in the surrounding space is recorded that their animal representations are implicitly maps of the relations of salient objects in their territories.
The basic structure of the animal system of representation can be seen by comparing the telesensory animal behavior guidance systems with that of the somatosensory animal. (See the accompanying diagrams of their animal behavior guidance systems.)
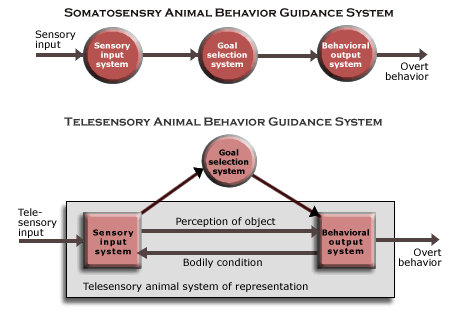
The animal behavior guidance system of somatosensory animals (and animals at lower biological levels) are so simple that their three subfunctions work together in the same way as the biological behavior guidance system (though they may be located throughout the animal body and recruit similar behavior in neighboring systems). Without an animal system of representation, they are unable to perceive objects at all.
In the telesensory animal behavior guidance system, what constitutes the animal system of representation is an interaction between mechanisms serving the sensory input and behavioral output subfunctions which is independent of the mechanisms serving the selection subfunction. An animal representation is constructed by combining telesensory input with input about the bodily condition. The bodily condition, such as which way the head and eyes are pointing, affects how telesensory input is registered, and how that input is registered is then used to determine how the animal moves, say, if it selects a goal that requires approaching the object. In telesensory animals, this mechanism is “hard wired” into the nervous system by the biological behavior guidance system in constructing it, and it is just a more or less elaborate set of rigid, instinctive routines that have been naturally selected for the power they afford.
The centralization of the nervous system is so obvious that no one denies the difference between "somatosensory" and "telesensory" nervous systems (though they many not use these terms). But the difference between them is not usually explained as a difference in level of neurological organization, and the main reason is that it is not recognized that there is a third level of neurological organization, much less a fourth. However, given that the function of the telesensory level of neurological organization is to provide an animal system of representation, however, we can see how higher levels of neurological organization could make animals even more powerful, including both spatio-temporal and structural imagination, because it is evident in the structure of their required subsystems. The following functional diagrams suggest their levels of part-whole complexity. These diagrams are explained more fully in relevant chapters, but even a gross impression of how they work shows how theses two higher levels of neurological organization would cause two step-like increases in animal power beyond telesensory animals..
Subjective animals. In subjective animals, the telesensory animal’s hard wired animal representation of the object is replaced by a kind of perception that is backed up by a faculty of imagination. The sensory input system still enables the animal to perceive the object in a way that guides locomotion in relation to it, but such perceptions are generated as part of a new interaction between the telesensory input and behavioral output systems, one in which covert locomotion in some direction can call up sequences of “local images” from memory representing the local scenes that would be encountered in moving in that direction (given a certain starting point).
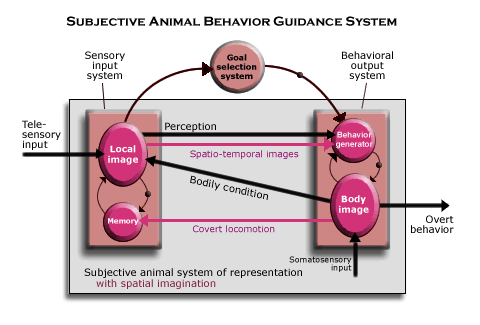
What makes such spatial imagination possible is, as this diagram suggests, a higher level of neurological organization in both the sensory input and behavioral output subsystems. (The higher level is indicated by the two circles within the enlarged square, representing the necessary subsystems.) Telesensory images are recorded in memory in sequences according to the locomotion that connects them (with memory using its own input from the current bodily condition), so that they can be called up in sequences from memory. (Since they are sequences of images over time, they are labeled “spatio-temporal images.”)
Once spatial imagination has been set up in the brain, therefore, to perceive an object is to be able to imagine how its spatial relations to objects would change, if the animal were to move its body in relation to it. And to imagine what would happen if the object were to move relative to its body and other objects. That is the effect of covert locomotion on subjective memory. To see the local image (and the objects it includes) as located within the map of its territory is to perceive the object as located in space. It also means perceiving its body as located in space, as we shall see. The subjective animal has, therefore, a conception of an object as located in space.
Manipulative animals. In the manipulative animal behavior guidance system, the sensory input system is still the animal’s perception of the object as located in (and potentially moving in) mammalian space. But more is perceived, because sensory input also represents the object as having a geometrical structure. Within the subjective animal representation of the object as located in space (the object in the local image which is part of its map of the territory), there is a further interaction between the sensory input and behavioral output systems in which covert manipulation can call up sequences of telesensory (and tactile) images of objects as they would appear if they were manipulated in certain ways. That yields an object image in the local image.
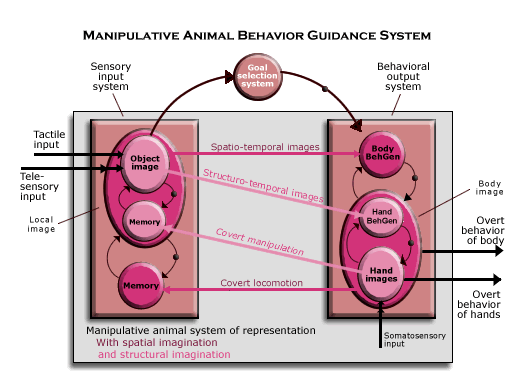
What makes structural imagination possible is, as the diagram suggests, a yet higher level of neurological organization in both the sensory input and behavior output subsystems (as indicated by the circles within the circles within the enlarged squares). (The connections representing perception and input from the current bodily condition, which are the underlying structure of the animal system of representation connecting the sensory input and behavioral output systems) are suppressed in this diagram for simplicity.) In manipulative animals, telesensory images of objects are recorded in memory in sequences along with tactile images from the hands according to how the object is being manipulated at the time (with memory using its own input from the current bodily condition), and these images can be called up in sequences from memory. (Since they are sequences of images over time, they are labeled “structuro-temporal images.”)
Once structural imagination has been set up in the brain, therefore, to perceive an object in space is not only to be able to imagine what would happen if it (or its own body) were to move around in space, but also what would happen if its hands were to manipulate the object in some way (for example, how it would appear if turned upside down or backside forward). That is to perceive the object in space as having a geometrical structure. Manipulative animals have, therefore, a conception of the object in space as having a geometrical structure.
The functions of sensory input and behavioral output systems, of the new subsystems, and then yet newer sub-subsystems that come to exist within them suggest the way in which higher levels of neurological organization can be functional in the sense of opening up, step by step, entire, new ranges of possible powers that animal behavior might evolve.
Starting with the telesensory stage, one new stage actualizes the potential power of spatial imagination, and after that, another new stage actualizes the potential power of structural imagination. Animal power is increased in each case by making the spatial aspects of animal behavior depend on spatial aspect of the world. The spatial aspects of the world they are exploiting are ontologically basic. In one case it is an intuitive understanding of spatial causation, and in the other it is an intuitive understanding of structural causation. These faculties of imagination approach a natural perfection for neurological mechanisms of their kind as part of the natural perfection of the multicellular animals to whose power they contribute, but since such powers are useful only in some ecological niches, they become most powerful only in some species of multicellular animals.
In each of the following three sections (telesensory animals, subjective animals, and manipulative animals), the function, evolution and structure of the nervous systems at each of their three levels of part-whole complexity are discussed in detail. Since higher levels of neurological organization can evolve only after gradual evolution has made mechanisms on the lower level of neurological organization reliable, if not perfect, in serving their functions, each level of neurological organization entails a stage of animal evolution in time, just as each level of biological organization entailed a stage of biological evolution. These are the animal stages of evolution.
Somatosensory animals do not represent a distinct stage of animal evolution, because they do not evolve after the evolution of multicellular animals, but as part of it. Simply having a nervous system does not put somatosensory animals on a higher level of organization than other multicellular organisms, because it is just a variety of multicellular organism that fills a kind of ecological niche, namely, acquiring free energy ingesting energy-rich objects. The structure of the somatosensory nervous system is basically just the structure of the body constructed by the mechanism of embryological development, because all three subsystems of the animal behavior guidance system are served locally by three different kinds of neurons at each location in the body: sensory neurons (sensory input), interneurons (goal selection), and motor neurons (behavioral output).
A higher level of neurological organization is required, as we have seen, for the animal to have an animal system of representation at all, and even in telesensory animals, single neurons can no longer serve the basic functions of registering telesensory input and generating behavioral output. Each such subfunction requires a whole group of neurons working together. That puts telesensory animals on a higher level of neurological organization than somatosensory animals, and as the diagrams suggest, higher levels of neurological organization involve higher levels of part-whole complexity within the mechanism serving the same basic subfunctions, sensory input and behavioral output. As we shall see more clearly when we consider the structure of the brain, the basic functions of generating perceptions and using them to guide behavior depend on two yet higher levels of neurological organization: circuits of groups neurons in subjective animals, and interactions among several similar circuits of neurons in manipulative animals.
Most animals, from worms and insects to reptiles and birds, are at the telesensory level of neurological organization. But the animal stages of evolution are somewhat more complicated than the stages of biological evolution, because there are two forms of embryological development and, thus, two series of levels/stages in multicellular animals. And the difference is relevant, because their biological behavior guidance systems use basically different mechanisms to organize groups of neurons to act as a whole within the nervous system. One uses ganglia, and the other uses two dimensional arrays of neurons laid out in a special neural tube. As a result there are two basically different kinds of telesensory animals, and only one of them is able to evolve beyond the telesensory stage. Invertebrates never evolve a subjective behavior guidance system, and we must explain why.
![]()